

Microbiota-driven antitumour immunity mediated by dendritic cell migration
- PMID: 40659786
- PMCID: PMC12390848
- DOI: 10.1038/s41586-025-09249-8
Microbiota-driven antitumour immunity mediated by dendritic cell migration
Abstract
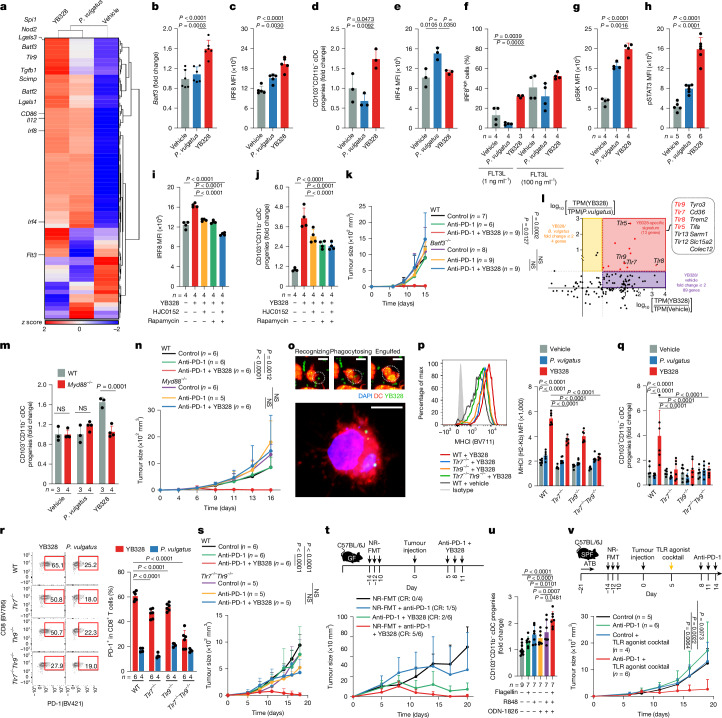
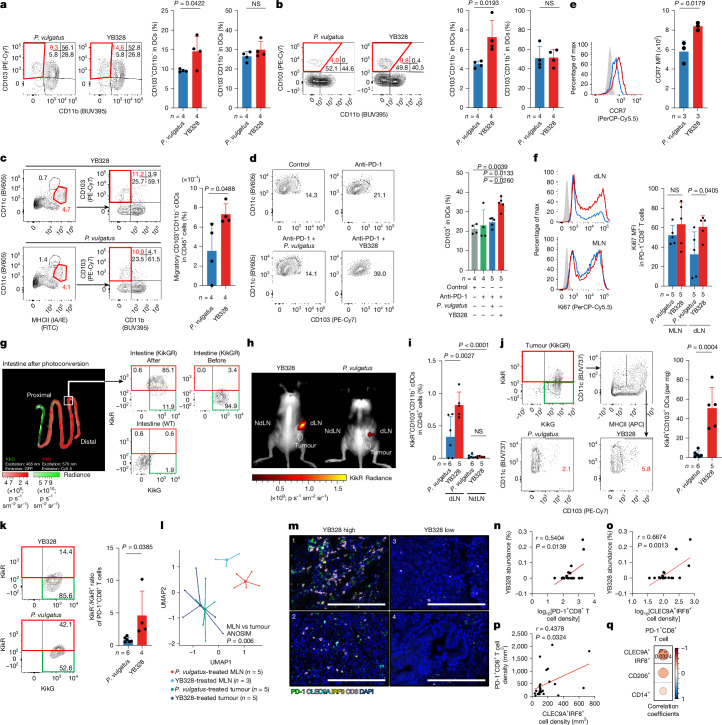
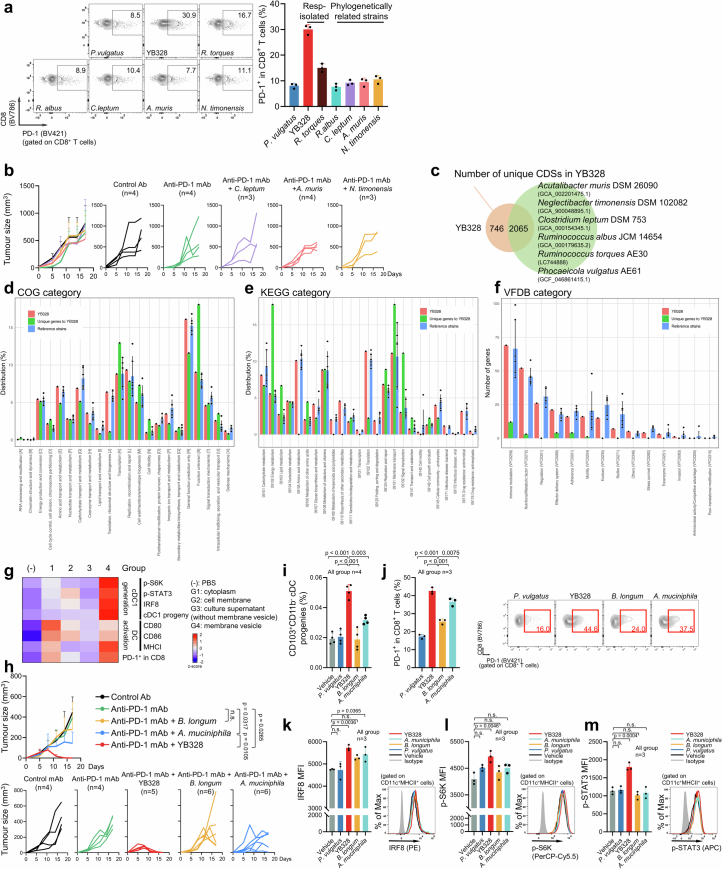
Gut microbiota influence the antitumour efficacy of immune checkpoint blockade1-6, but the mechanisms of action have not been fully elucidated. Here, we show that a new strain of the bacterial genus Hominenteromicrobium (designated YB328) isolated from the faeces of patients who responded to programmed cell death 1 (PD-1) blockade augmented antitumour responses in mice. YB328 activated tumour-specific CD8+ T cells through the stimulation of CD103+CD11b- conventional dendritic cells (cDCs), which, following exposure in the gut, migrated to the tumour microenvironment. Mice showed improved antitumour efficacy of PD-1 blockade when treated with faecal transplants from non-responder patients supplemented with YB238. This result suggests that YB328 could function in a dominant manner. YB328-activated CD103+CD11b- cDCs showed prolonged engagement with tumour-specific CD8+ T cells and promoted PD-1 expression in these cells. Moreover, YB238-augmented antitumour efficacy of PD-1 blockade treatment was observed in multiple mouse models of cancer. Patients with elevated YB328 abundance had increased infiltration of CD103+CD11b- cDCs in tumours and had a favourable response to PD-1 blockade therapy in various cancer types. We propose that gut microbiota enhance antitumour immunity by accelerating the maturation and migration of CD103+CD11b- cDCs to increase the number of CD8+ T cells that respond to diverse tumour antigens.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: S. Koyama receives research funding from Otsuka Pharmaceutical and Chugai Pharmaceutical outside this study and receives honoraria from MSD and Chugai Pharmaceutical. K.I. receives honoraria from MSD and Chugai Pharmaceutical. M.N. receives honoraria (lecture fees) from MSD, Ono Pharmaceutical and Bristol-Myers Squibb (BMS). A.K. receives personal fees from Daiichi-Sankyo, Lilly, Ono Pharmaceutical, Taiho Pharmaceutical, BMS, Merck Pharmaceutical, Sumitomo Dainippon Pharma and AstraZeneca outside this study. K.W. serves as a board member and a founder of ARC Therapies outside of this study. T.D. receives personal fees for advisory roles from Sumitomo Dainippon Pharma, Taiho Pharmaceutical, Takeda Pharmaceutical, Chugai Pharmaceutical, AbbVie, Bayer, Rakuten Medical, Otsuka Pharmaceutical, KAKEN Pharmaceutical, Kyowa Kirin, SHIONOGI, PRA Health Science, A2 Health Care, Noile-Immune Biotech, MSD, Daiichi-Sankyo, Amgen, Novartis, Boehringer Ingelheim, Janssen Pharmaceutical and Astellas Pharmaceutical; receives honoraria (lecture fees) from BMS, Rakuten Medical, Ono Pharmaceutical, Daiichi-Sankyo and AstraZeneca; and receives research funding from Lilly, MSD, Daiichi-Sankyo, Sumitomo Dainippon Pharma, Taiho Pharmaceutical, Novartis, Merck Pharmaceutical, Janssen Pharmaceutical, Boehringer Ingelheim, Pfizer, BMS, AbbVie, Eisai, IQVIA, Chugai Pharmaceutical and SHIONOGI outside this study. K.S. reports receiving personal fees for consulting and advisory roles from BMS, Takeda, Ono Pharmaceutical, Novartis, Daiichi Sankyo, Amgen, Boehringer Ingelheim, Merck Pharmaceutical, Astellas, Guardant Health Japan, Janssen, AstraZeneca, Zymeworks Biopharmaceuticals, ALX Oncology and Bayer; receiving honoraria from BMS, Ono Pharmaceutical, Janssen, Eli Lilly, Astellas and AstraZeneca; and receiving research funding (all to the institution) from Astellas, Ono Pharmaceutical, Daiichi Sankyo, Taiho Pharmaceutical, Chugai, Merck Pharmaceutical, Amgen, Eisai, PRA Health Sciences and Syneos Health, outside the submitted work. H.M. receives research grants from Ono Pharmaceutical, Daiichi Sankyo, PFDeNA, Konica-Minolta and Ambry Genetics and serves as a board member of CureGene outside this study. H.N. receives research funding and honoraria (lecture fees) from Ono Pharmaceutical, BMS, Chugai Pharmaceutical, BD Japan and MSD; receives honoraria (lecture fees) from Amgen; receives research funding from Taiho Pharmaceutical, Daiichi-Sankyo, Kyowa Kirin, Zenyaku Kogyo, Oncolys BioPharma, Debiopharma, Asahi-Kasei, Sysmex, Fujifilm, SRL, Astellas Pharmaceutical, Sumitomo Dainippon Pharma, ARC Therapies and RIKAKEN Holdings; and serves as a scientific advisor and a founder of ARC Therapies and a scientific advisor of LTZ Therapeutics outside this study. S.F., Y.B. and H.N. are the primary inventors on pending patents 2020-165470 belonging to RIKEN and the National Cancer Center Japan. The other authors declare no competing interests.
Figures














References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
