

An expanded subventricular zone supports postnatal cortical interneuron migration in gyrencephalic brains
- PMID: 40659844
- PMCID: PMC12321571
- DOI: 10.1038/s41593-025-01987-2
An expanded subventricular zone supports postnatal cortical interneuron migration in gyrencephalic brains
Abstract
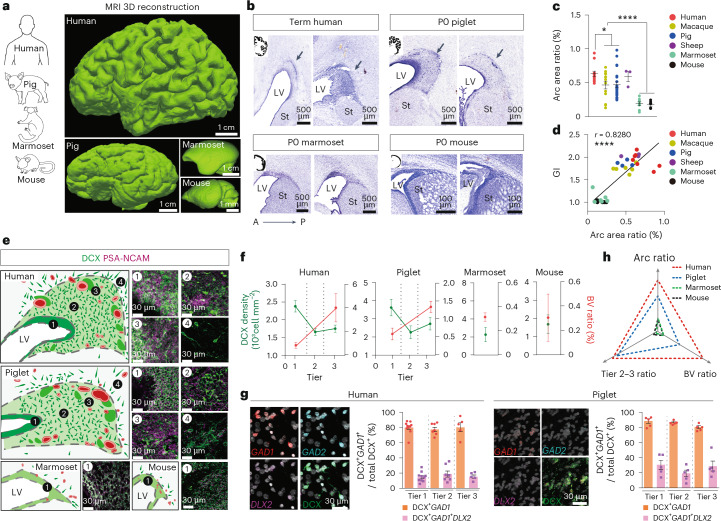
Cortical GABAergic interneurons generated in the ventral developing brain travel long distances to their final destinations. While there are examples of interneuron migration in the neonatal human brain, the extent of postnatal migration across species and how it contributes to cortical interneuron composition remains unknown. Here we demonstrate that neonatal gyrencephalic brains, including humans, nonhuman primates and piglets, harbor an elaborate subventricular zone, termed the Arc, due to its curved morphology and expanded neuroblast populations. The Arc is absent in lissencephalic marmoset and mouse brains. Transcriptomic and histological approaches revealed that Arc neurons are diverse interneurons from the medial and caudal ganglionic eminences that migrate into the frontal, cingulate and temporal cortex. Arc-cortical targets exhibit an increase in VIP+ neuronal density compared to other regions. Our findings reveal that the Arc is a developmental structure that supports the expansion of postnatal neuronal migration for cortical interneuron patterning in gyrencephalic brains.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures














References
-
- Ayala, R., Shu, T. & Tsai, L. H. Trekking across the brain: the journey of neuronal migration. Cell128, 29–43 (2007). - PubMed
-
- Buchsbaum, I. Y. & Cappello, S. Neuronal migration in the CNS during development and disease: insights from in vivo and in vitro models. Development146, dev163766 (2019). - PubMed
-
- Morshead, C. M. et al. Neural stem cells in the adult mammalian forebrain: a relatively quiescent subpopulation of subependymal cells. Neuron13, 1071–1082 (1994). - PubMed
-
- Gage, F. H. Mammalian neural stem cells. Science287, 1433–1438 (2000). - PubMed
MeSH terms
Grants and funding
- R24 NS092988/NS/NINDS NIH HHS/United States
- P01 NS083513/NS/NINDS NIH HHS/United States
- DP2 NS122550/NS/NINDS NIH HHS/United States
- NS083513, NS122550-01/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
