

This is a preprint.
Oncogenic and tumor-suppressive forces converge on a progenitor-orchestrated niche to shape early tumorigenesis
- PMID: 40661354
- PMCID: PMC12259082
- DOI: 10.1101/2025.06.10.656791
Oncogenic and tumor-suppressive forces converge on a progenitor-orchestrated niche to shape early tumorigenesis
Abstract
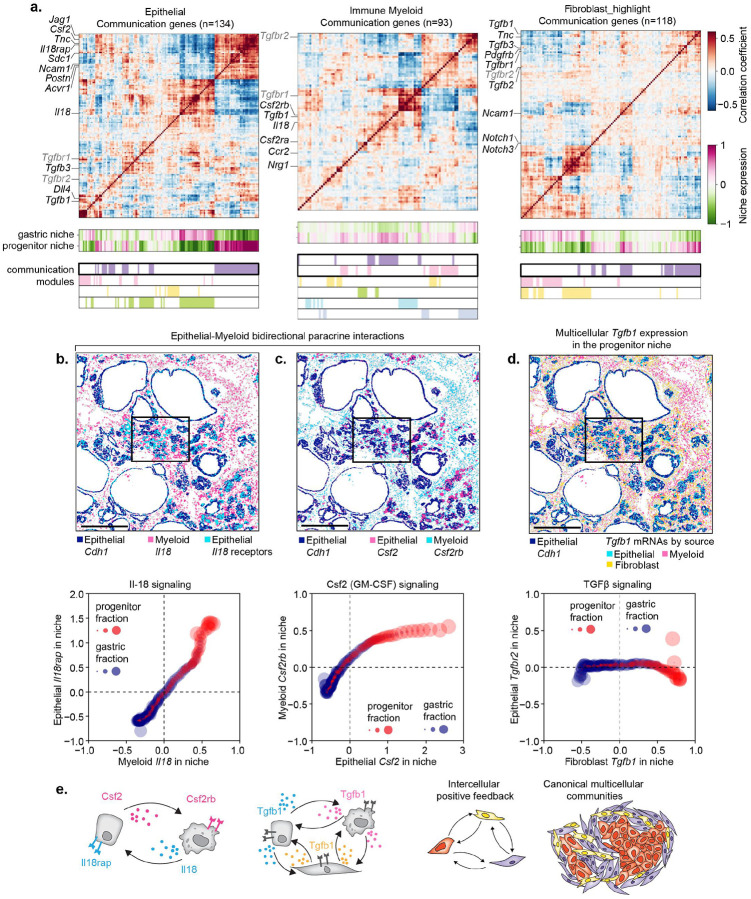
The transition from benign to malignant growth is a pivotal yet poorly understood step in cancer progression that marks the shift from a pathologically inert condition to a clinically lethal disease. Here, we integrate lineage tracing, single-cell and spatial transcriptomics to visualize the molecular, cellular and tissue-level events that promote or restrain malignancy during the tumor initiation in mouse models of pancreatic ductal adenocarcinoma (PDAC). We identify a discrete progenitor-like population of KRAS-mutant cells that co-activates oncogenic and tumor-suppressive programs-including p53, CDKN2A, and SMAD4-engaging senescence-like responses and remodeling their microenvironment, ultimately assembling a niche that mirrors invasive PDAC. KRAS inhibition depletes progenitor-like cells and dismantles their niche. Conversely, p53 suppression enables progenitor cell expansion, epithelial-mesenchymal reprogramming, and immune-privileged niche formation. These findings position the progenitor-like state as the convergence point of cancer-driving mutations, plasticity, and tissue remodeling-revealing a critical window for intercepting malignancy at its origin.
Conflict of interest statement
DECLARATION OF INTERESTS While not directly related to this manuscript, S.W.L. declares competing interests outside consultancy and equity for Oric Pharmaceuticals, Blueprint Medicines, Mirimus, Senecea Therapeutics, Faeth Therapeutics and PMV Pharmaceuticals; and outside consultancy (no equity) for Fate Therapeutics. D.P. reports equity interests and provision of services for Insitro, Inc. P.B.R provides compensated professional services and activities for EMD Serono, Faeth Therapeutics, Urogen Pharma, Incyte, and Natera Inc. He also offers uncompensated professional services and activities for 10x Genomics, XRad Therapeutics, and the HPV Alliance and Anal Cancer Foundation non-profit organizations. The other authors declare no competing interests.
Figures






References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous