

This is a preprint.
Structural Identification of Major Molecular Determinants for Phosphotyrosine Recognition in Tyrosine Kinases Reveals Tumour Promoting and Suppressive Functions
- PMID: 40661594
- PMCID: PMC12259089
- DOI: 10.1101/2025.06.10.658871
Structural Identification of Major Molecular Determinants for Phosphotyrosine Recognition in Tyrosine Kinases Reveals Tumour Promoting and Suppressive Functions
Abstract
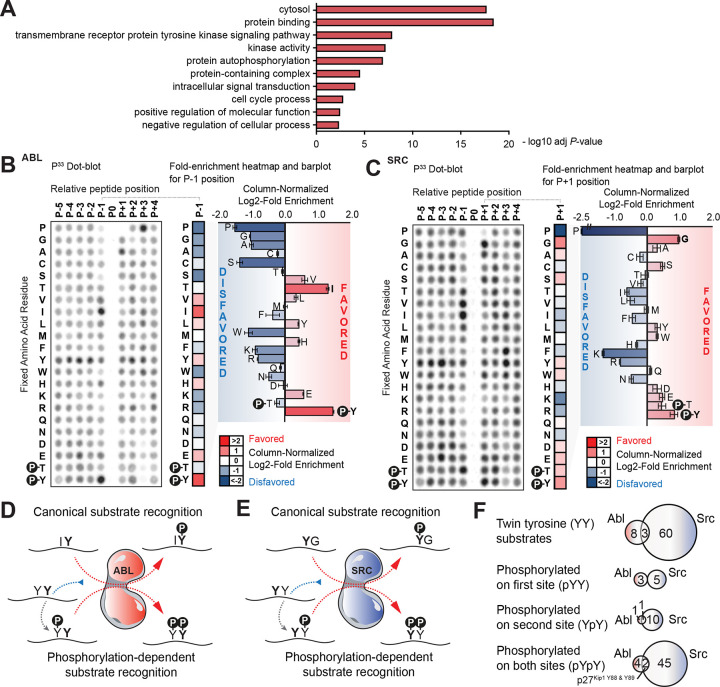
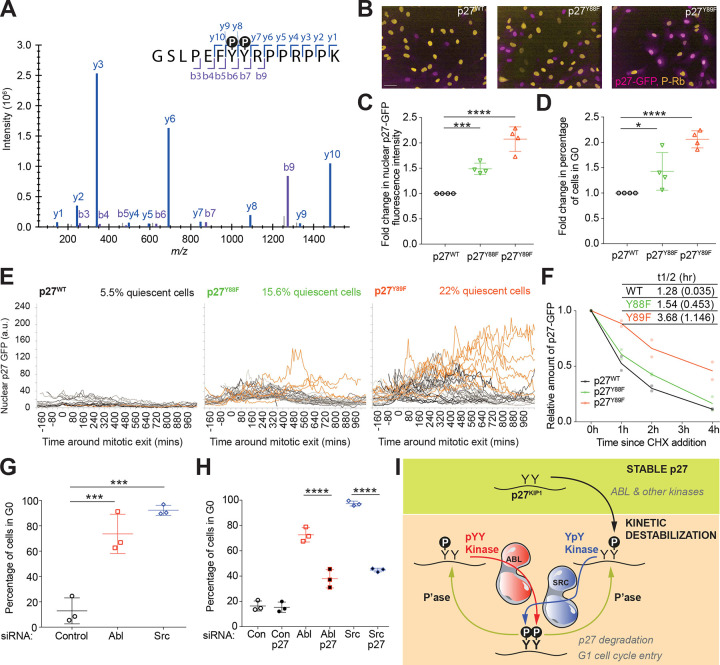
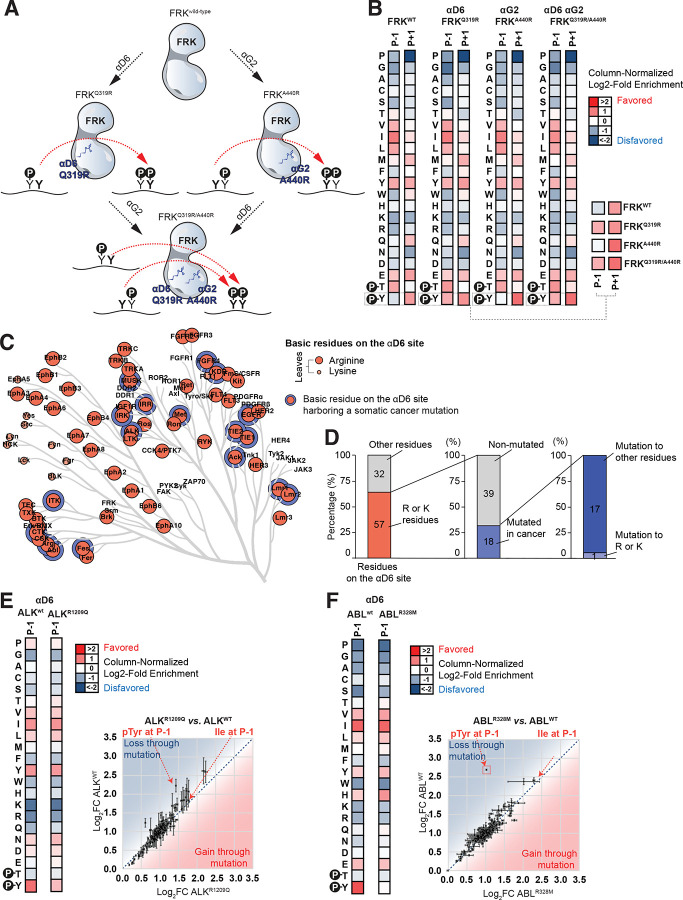
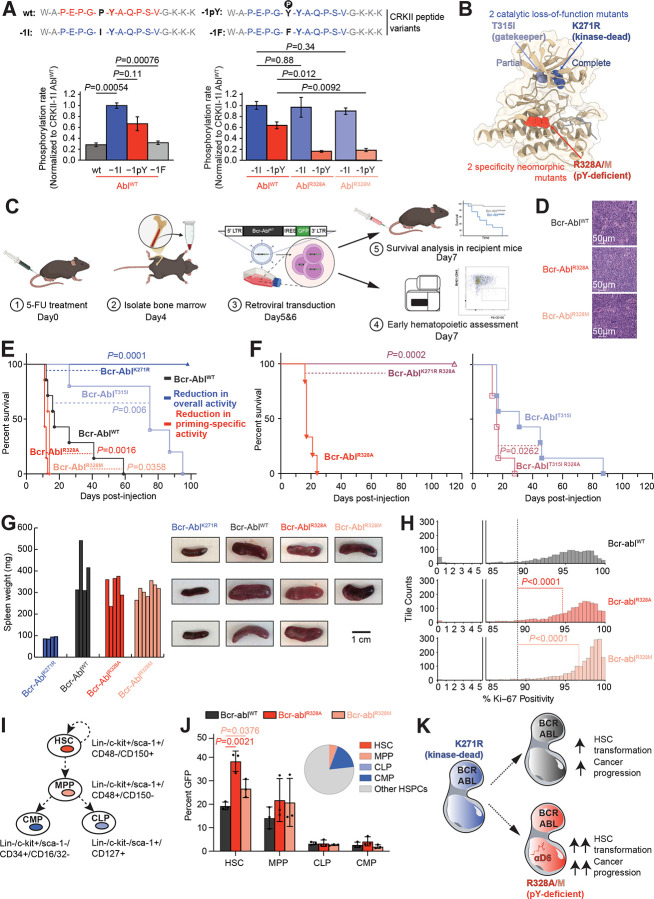
Protein tyrosine kinases activate signaling pathways by catalyzing the phosphorylation of tyrosine residues in their substrates. Mounting evidence suggests that, in addition to recognizing phosphorylated tyrosine (pTyr) residues through specific phosphobinding modules, many protein kinases selectively recognize pTyr directly adjacent to the tyrosine residue they phosphorylate and catalyze the formation of twin pTyr-pTyr sites. Here, we demonstrate the importance of this phosphopriming-driven twin pTyr signaling in promoting cell cycle progression through the cell cycle-inhibitory protein p27Kip1. We identify, structurally resolve, and tune two distinct molecular determinants driving the selective recognition of pTyr directly N- and C-terminal to the target phospho-acceptor tyrosine site. We further show structural and biochemical conservation in this recognition, and identify cancer-associated alterations to these determinants that are unable to recognize phosphoprimed substrates. Finally, using an in vivo mouse model of leukemia we show that Bcr-Abl mutants unable to recognize phosphoprimed substrates paradoxically result in enhanced tumor development and progression. These data indicate that Bcr-Abl, like other proto-oncogenes such as Ras or Myc, engages both pro- and anti-oncogenic programs - but in the case of Bcr-Abl, this is accomplished through a mechanism involving traditional and phosphoprimed substrate recognition.
Conflict of interest statement
L.C.C. is a founder and member of the board of directors of Agios Pharmaceuticals and is a founder and receives research support from Petra Pharmaceuticals; is listed as an inventor on a patent (WO2019232403A1, Weill Cornell Medicine) for combination therapy for PI3K-associated disease or disorder, and the identification of therapeutic interventions to improve response to PI3K inhibitors for cancer treatment; is a co-founder and shareholder in Faeth Therapeutics; has equity in and consults for Cell Signaling Technologies, Volastra, Larkspur and 1 Base Pharmaceuticals; and consults for Loxo-Lilly. J.L.J has received consulting fees from Scorpion Therapeutics and Volastra Therapeutics. M.B.Y. is on the Scientific Advisory Board of Odyssey Therapeutics. The remaining authors declare no competing interests.
Figures



References
-
- Eckhart W., Hutchinson M. A. & Hunter T. An activity phosphorylating tyrosine in polyoma T antigen immunoprecipitates. Cell 18, 925–933 (1979). - PubMed
-
- Smith A. E., Smith R., Griffin B. & Fried M. Protein kinase activity associated with polyoma virus middle T antigen in vitro. Cell 18, 915–924 (1979). - PubMed
-
- Schaffhausen B. S. & Benjamin T. L. Phosphorylation of polyoma T antigens. Cell 18, 935–946 (1979). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous