

This is a preprint.
NDR kinase SAX-1 controls dendrite branch-specific elimination during neuronal remodeling in C. elegans
- PMID: 40661618
- PMCID: PMC12259037
- DOI: 10.1101/2025.06.09.658633
NDR kinase SAX-1 controls dendrite branch-specific elimination during neuronal remodeling in C. elegans
Abstract
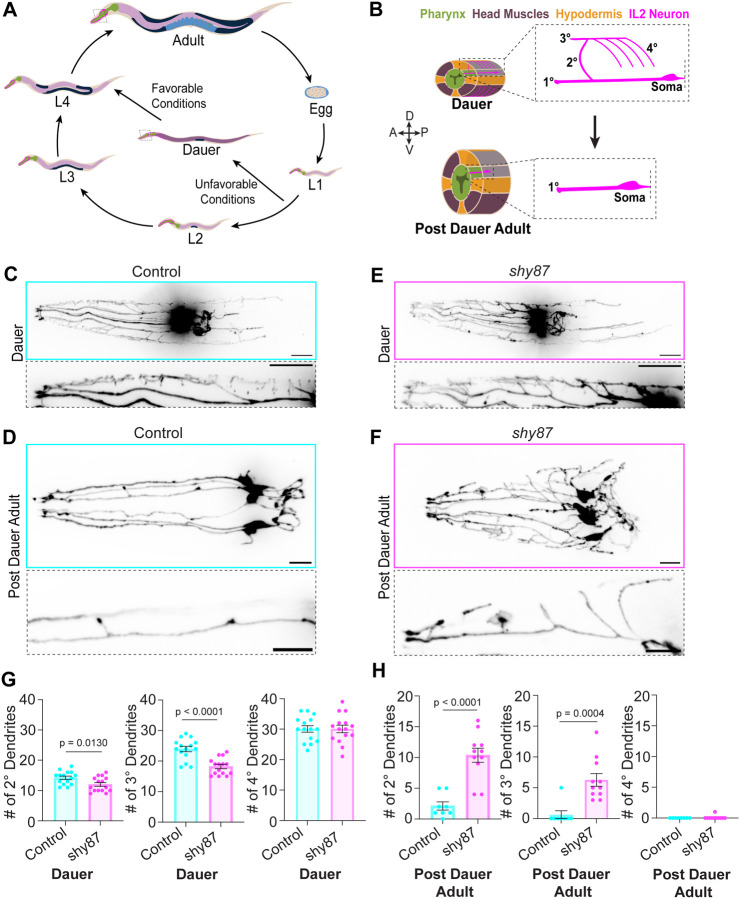
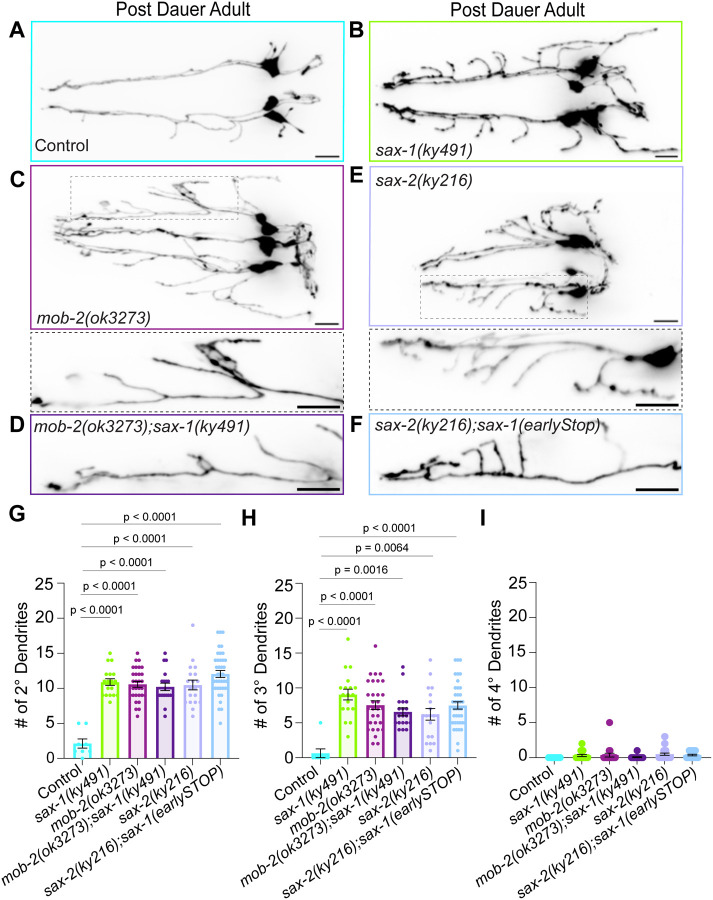
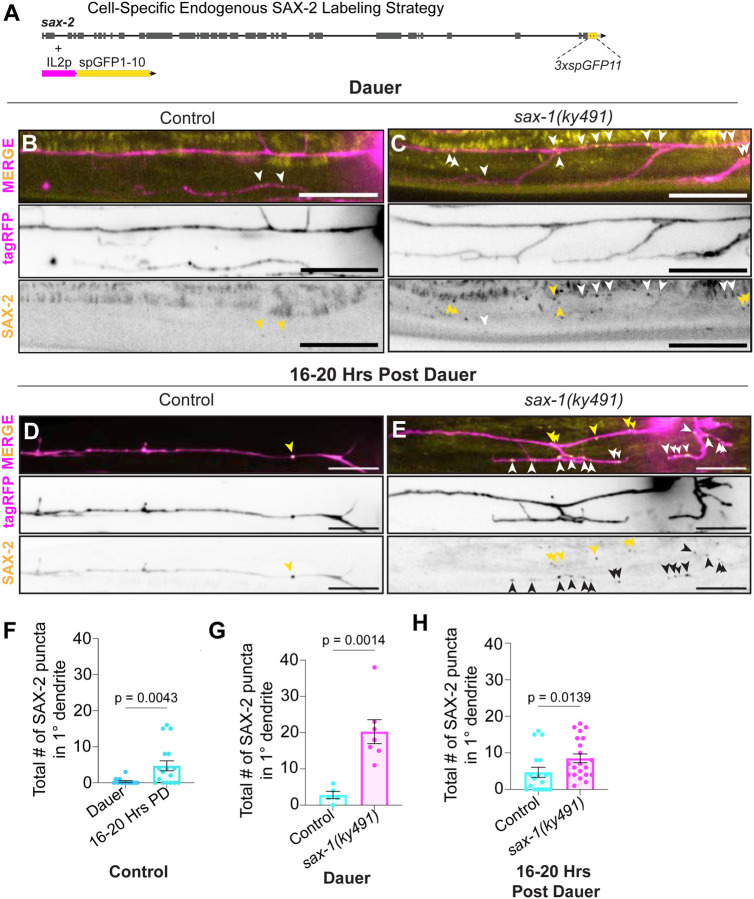
Neuronal remodeling is crucial for proper nervous system development and function, and can be initiated by developmental programs, activity-dependent mechanisms, or stress. Despite significant advances, the underlying mechanisms that govern this process remain poorly understood. Here, we adapted C. elegans IL2 sensory dendrites as a model system to study developmental and stress-mediated dendrite pruning. Upon entering a stress-induced developmental diapause, IL2 dendrites grow a complex dendritic arbor, which is later pruned when reproductive development resumes. We identified unexpected specificity in the pruning process, with distinct genetic requirements to direct branch-specific elimination of secondary, tertiary, and quaternary branches. The serine/threonine kinase SAX-1/NDR promotes elimination of secondary and tertiary, but not quaternary, dendrites. SAX-1 functions with its conserved interactors SAX-2/Furry and MOB-2 in the removal of both dendritic branches. The guanine-nucleotide exchange factor RABI-1/Rabin8 and the small GTPase RAB-11.2 mediate the elimination of secondary branches with SAX-1, but their effect on tertiary branches is minimal. Consistent with the known roles of RABI-1 and RAB-11.2 in regulating membrane dynamics, we find that SAX-1 promotes endocytosis during remodeling. Together, our findings reveal distinct mechanisms for branch-specific elimination under stress-induced and developmentally regulated neuronal remodeling.
Keywords: MOB-2/Mob; RABI-1/Rabin8; SAX-1/NDR; SAX-2/Fry; dauer; dendrite pruning; membrane trafficking; neuronal remodeling; small GTPase RAB-11.2.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures



Similar articles
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
RABR-1, an atypical Rab-related GTPase, cell-nonautonomously restricts somatosensory dendrite branching.Genetics. 2024 Oct 7;228(2):iyae113. doi: 10.1093/genetics/iyae113. Genetics. 2024. PMID: 39028768 Free PMC article.
-
AMIGO1 Promotes Axon Growth and Territory Matching in the Retina.J Neurosci. 2022 Mar 30;42(13):2678-2689. doi: 10.1523/JNEUROSCI.1164-21.2022. Epub 2022 Feb 15. J Neurosci. 2022. PMID: 35169021 Free PMC article.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
EORTC guidelines for the use of erythropoietic proteins in anaemic patients with cancer: 2006 update.Eur J Cancer. 2007 Jan;43(2):258-70. doi: 10.1016/j.ejca.2006.10.014. Epub 2006 Dec 19. Eur J Cancer. 2007. PMID: 17182241
References
-
- Altun Z.F. and Hall D.H. 2009. “Introduction.” WormAtlas. 10.3908/wormatlas.1.1 - DOI
-
- Brunson Kristen L., Enikö Kramár, Lin Bin, Chen Yuncai, Colgin Laura Lee, Yanagihara Theodore K., Lynch Gary, and Baram Tallie Z.. 2005. “Mechanisms of Late-Onset Cognitive Decline after Early-Life Stress.” Journal of Neuroscience 25 (41): 9328–38. 10.1523/JNEUROSCI.228-105.2005. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous