

EPHA4 signaling dysregulation links abnormal locomotion and the development of idiopathic scoliosis
- PMID: 40662934
- PMCID: PMC12263152
- DOI: 10.7554/eLife.95324
EPHA4 signaling dysregulation links abnormal locomotion and the development of idiopathic scoliosis
Abstract
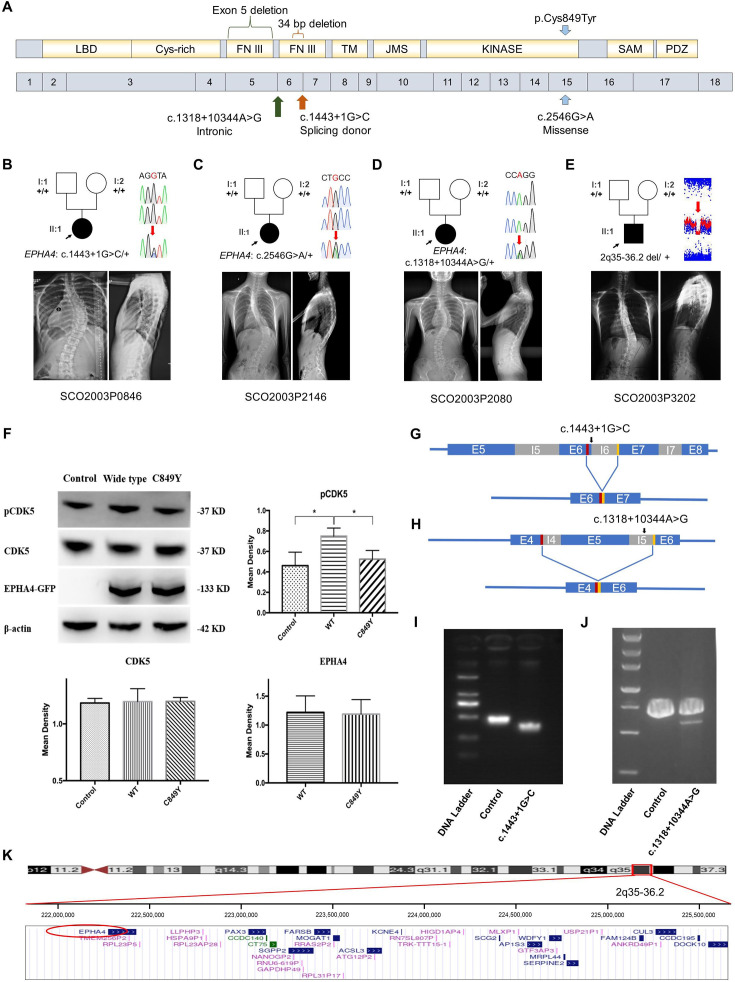
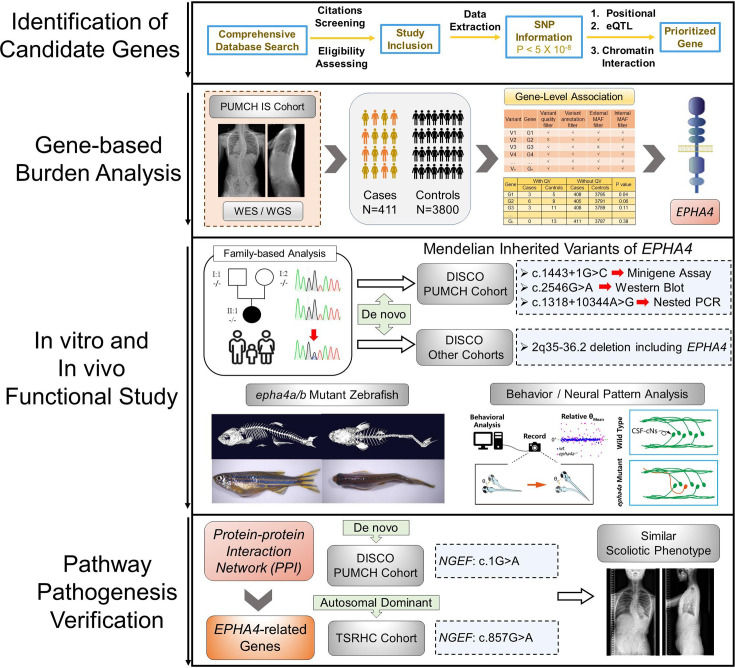
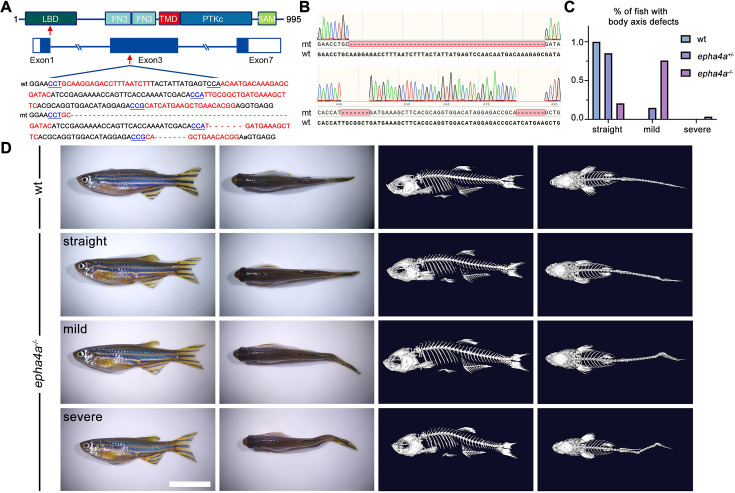
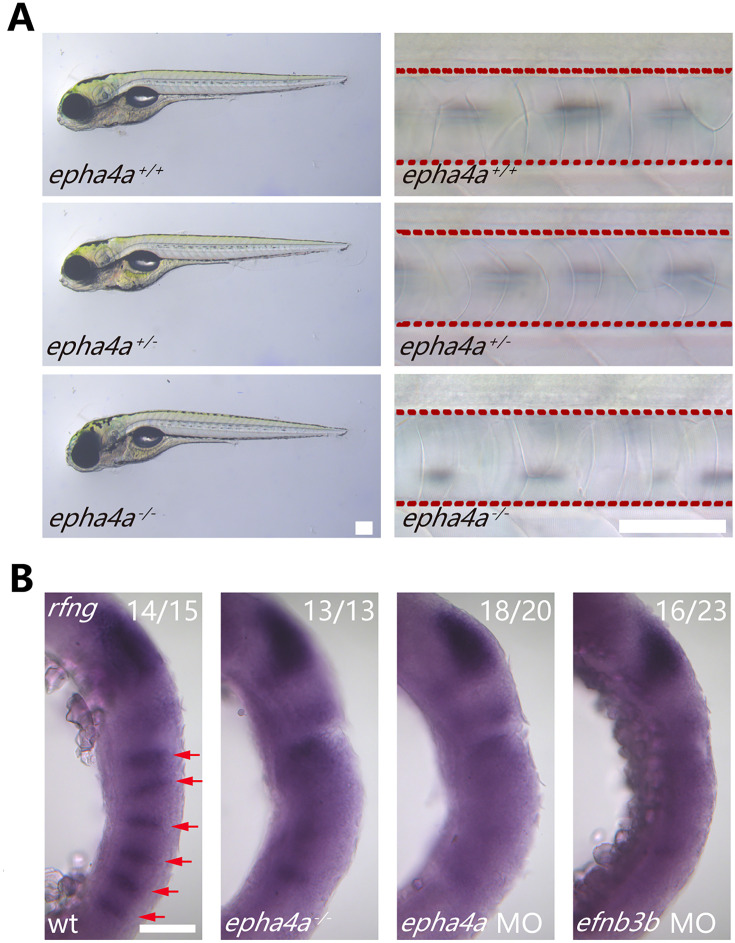
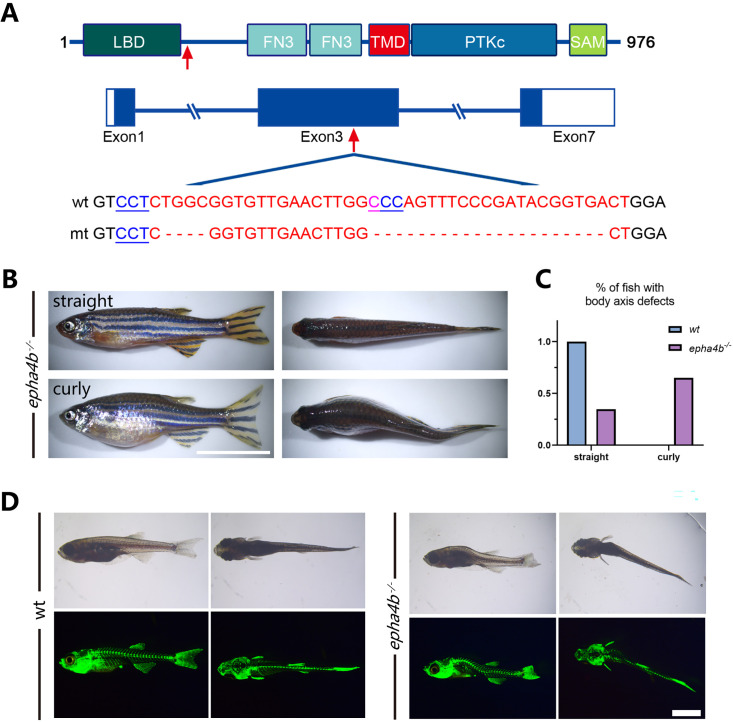
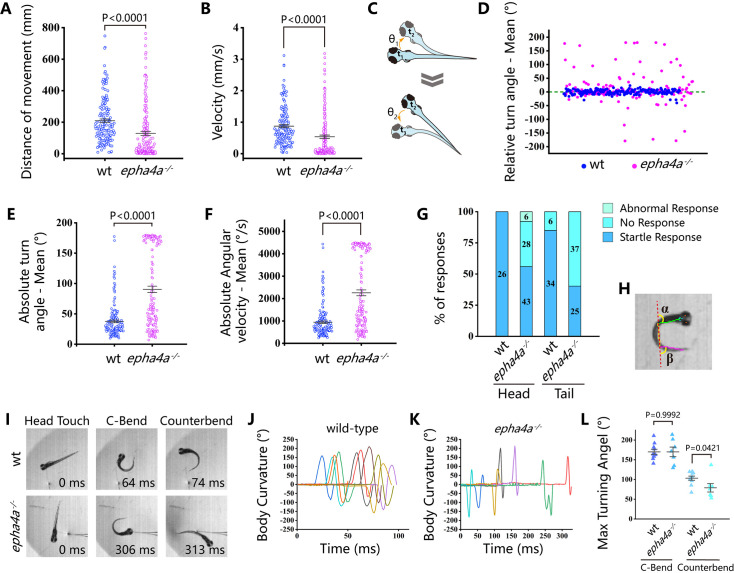
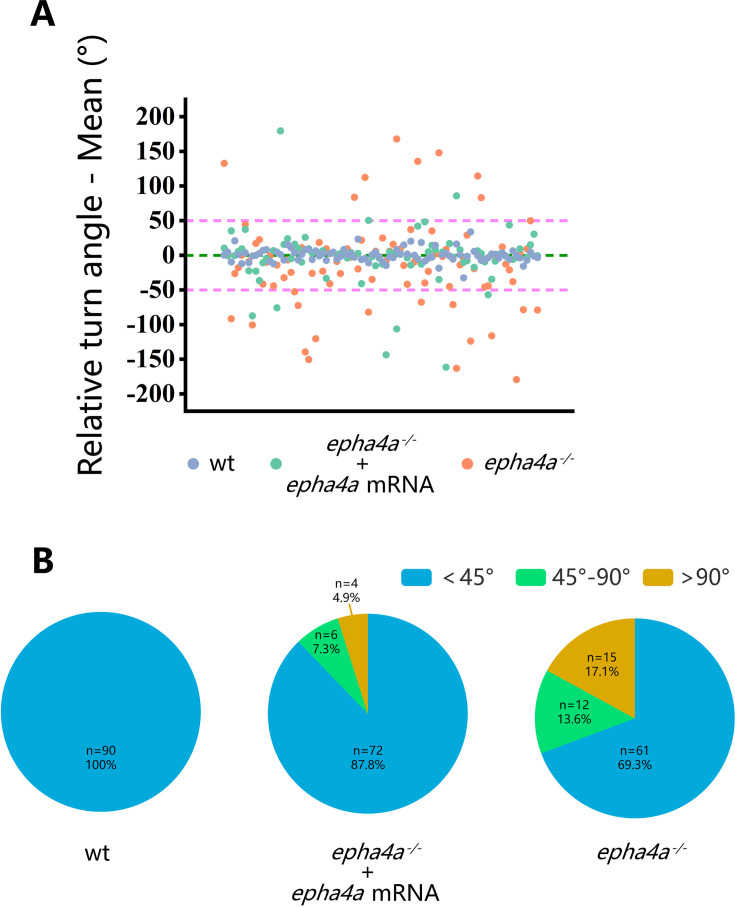
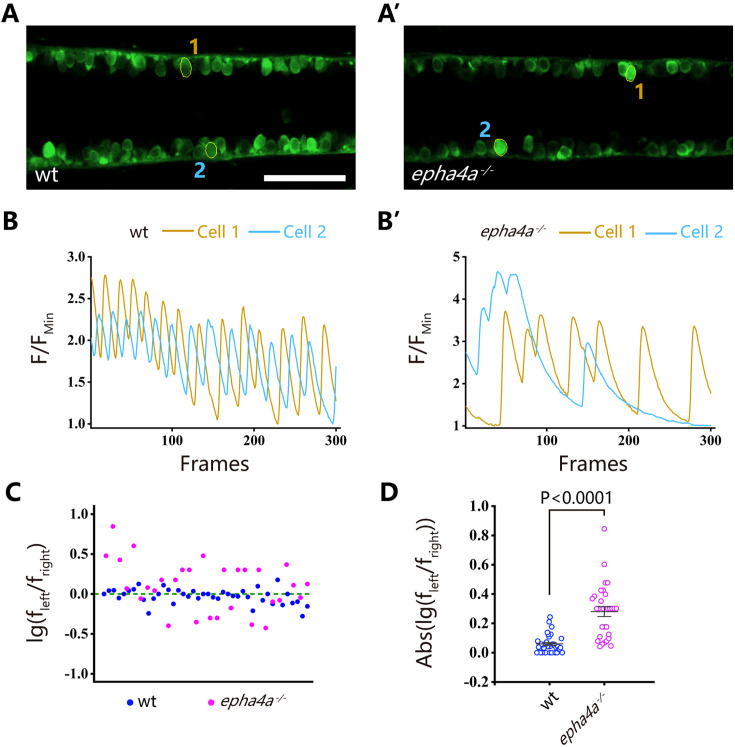
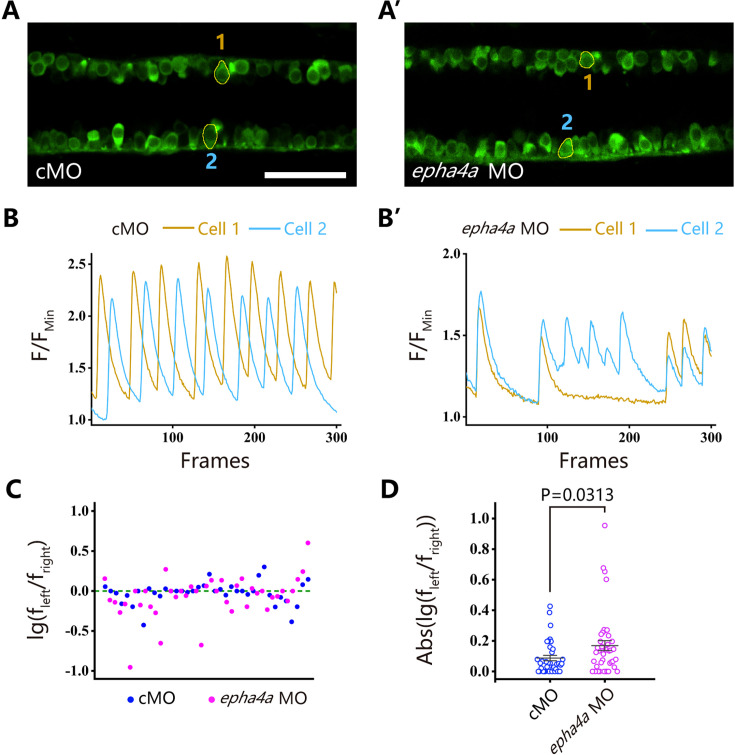
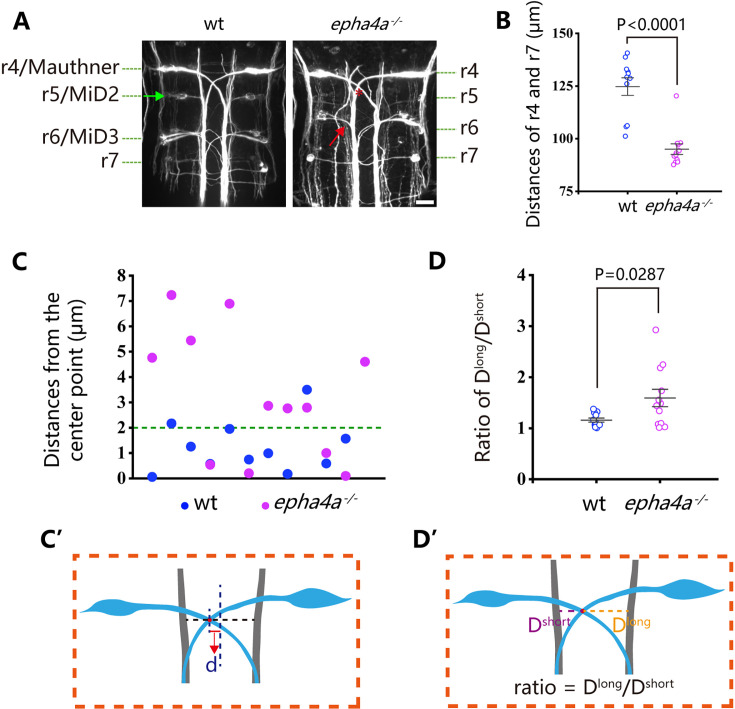
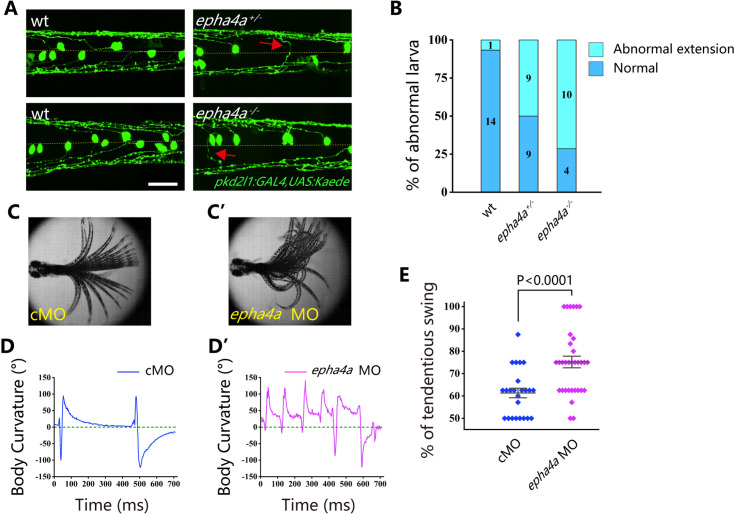
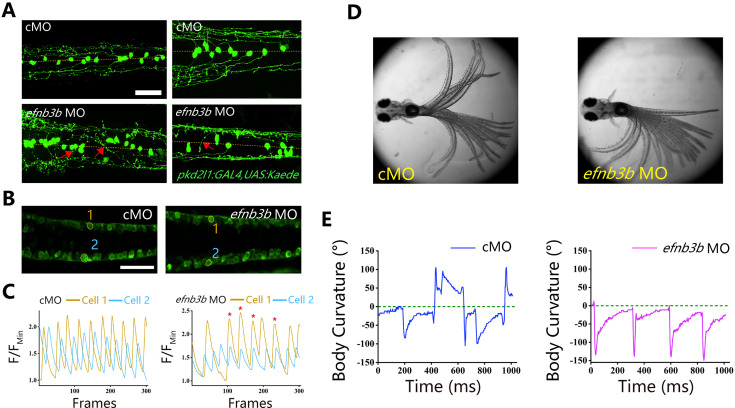
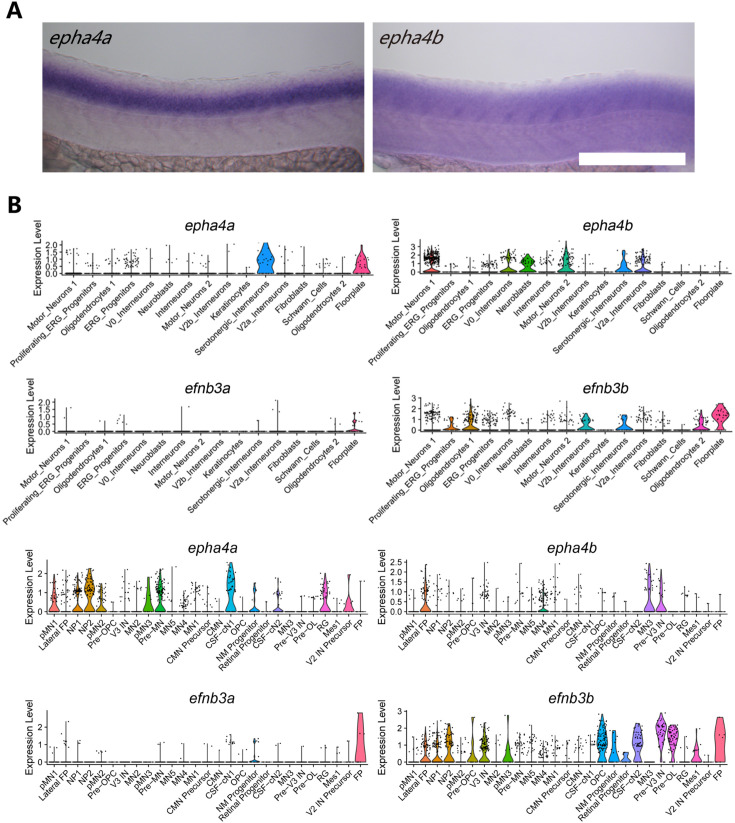
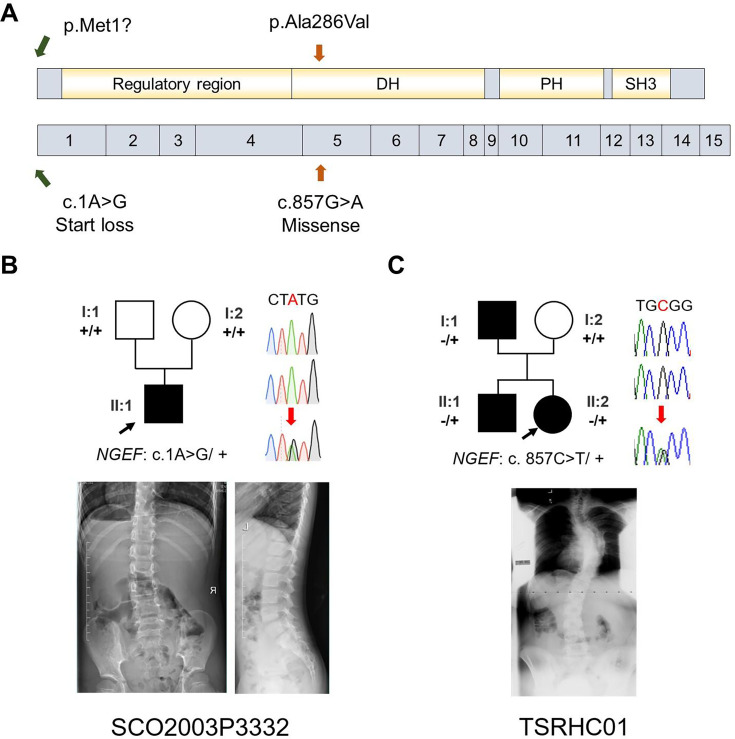
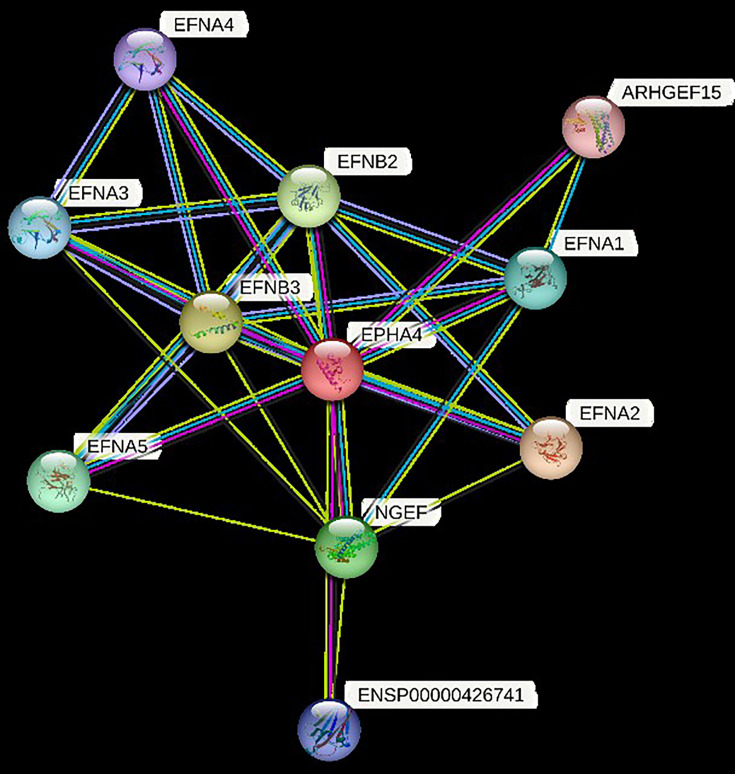
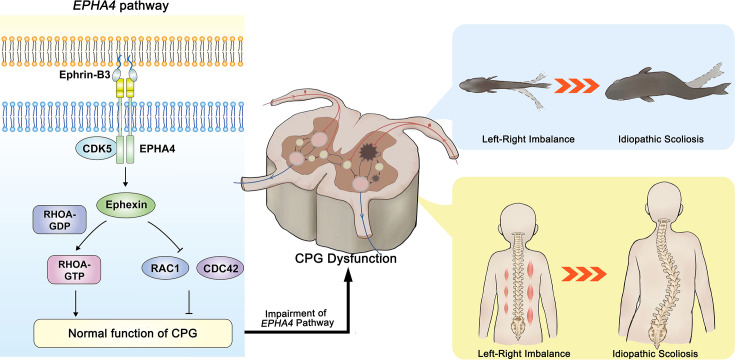
Idiopathic scoliosis (IS) is the most common form of spinal deformity with unclear pathogenesis. In this study, we first reanalyzed the loci associated with IS, drawing upon previous studies. Subsequently, we mapped these loci to candidate genes using either location-based or function-based strategies. To further substantiate our findings, we verified the enrichment of variants within these candidate genes across several large IS cohorts encompassing Chinese, East Asian, and European populations. Consequently, we identified variants in the EPHA4 gene as compelling candidates for IS. To confirm their pathogenicity, we generated zebrafish mutants of epha4a. Remarkably, the zebrafish epha4a mutants exhibited pronounced scoliosis during later stages of development, effectively recapitulating the IS phenotype. We observed that the epha4a mutants displayed defects in left-right coordination during locomotion, which arose from disorganized neural activation in these mutants. Our subsequent experiments indicated that the disruption of the central pattern generator (CPG) network, characterized by abnormal axon guidance of spinal cord interneurons, contributed to the disorganization observed in the mutants. Moreover, when knocked down efnb3b, the ligand for Epha4a, we observed similar CPG defects and disrupted left-right locomotion. These findings suggested that ephrin B3-Epha4 signaling is vital for the proper functioning of CPGs, and defects in this pathway could lead to scoliosis in zebrafish. Furthermore, we identified two cases of IS in NGEF, a downstream molecule in the EPHA4 pathway. Collectively, our data provide compelling evidence that neural patterning impairments and disruptions in CPGs may underlie the pathogenesis of IS.
Keywords: EPHA4; central pattern generators; genetics; genomics; human; idiopathic scoliosis; medicine; pathogenesis; zebrafish.
© 2024, Wang, Yang, Zhao et al.
Conflict of interest statement
LW, XY, SZ, PZ, WW, KX, XC, QL, AK, YK, JL, XF, NO, ZC, YL, LL, HX, PZ, XL, YN, SW, SL, SY, CT, ZL, SC, XZ, ZW, GQ, JL, JR, CW, JZ, CZ, NW No competing interests declared, PL, JP affiliated to Baylor Genetics, Houston, SI J.R.L has stock ownership in 23andMe, is a paid consultant for Regeneron Pharmaceuticals and Novartis, and is a co-inventor on multiple United States and European patents related to molecular diagnostics for inherited neuropathies, eye diseases and bacterial genomic fingerprinting
Figures

Update of
- doi: 10.1101/2024.03.04.583361
- doi: 10.7554/eLife.95324.1
- doi: 10.7554/eLife.95324.2
References
-
- Aguet F, Anand S, Ardlie KG, Gabriel S, Getz GA, Graubert A, Hadley K, Handsaker RE, Huang KH, Kashin S, Li X, MacArthur DG, Meier SR, Nedzel JL, Nguyen DT, Segrè AV, Todres E, Balliu B, Barbeira AN, Battle A, Bonazzola R, Brown A, Brown CD, Castel SE, Conrad DF, Cotter DJ, Cox N, Das S, de Goede OM, Dermitzakis ET, Einson J, Engelhardt BE, Eskin E, Eulalio TY, Ferraro NM, Flynn ED, Fresard L, Gamazon ER, Garrido-Martín D, Gay NR, Gloudemans MJ, Guigó R, Hame AR, He Y, Hoffman PJ, Hormozdiari F, Hou L, Im HK, Jo B, Kasela S, Kellis M, Kim-Hellmuth S, Kwong A, Lappalainen T, Li X, Liang Y, Mangul S, Mohammadi P, Montgomery SB, Muñoz-Aguirre M, Nachun DC, Nobel AB, Oliva M, Park Y, Park Y, Parsana P, Rao AS, Reverter F, Rouhana JM, Sabatti C, Saha A, Stephens M, Stranger BE, Strober BJ, Teran NA, Viñuela A, Wang G, Wen X, Wright F, Wucher V, Zou Y, Ferreira PG, Li G, Melé M, Yeger-Lotem E, Barcus ME, Bradbury D, Krubit T, McLean JA, Qi L, Robinson K, Roche NV, Smith AM, Sobin L, Tabor DE, Undale A, Bridge J, Brigham LE, Foster BA, Gillard BM, Hasz R, Hunter M, Johns C, Johnson M, Karasik E, Kopen G, Leinweber WF, McDonald A, Moser MT, Myer K, Ramsey KD, Roe B, Shad S, Thomas JA, Walters G, Washington M, Wheeler J, Jewell SD, Rohrer DC, Valley DR, Davis DA, Mash DC, Branton PA, Barker LK, Gardiner HM, Mosavel M, Siminoff LA, Flicek P, Haeussler M, Juettemann T, Kent WJ, Lee CM, Powell CC, Rosenbloom KR, Ruffier M, Sheppard D, Taylor K, Trevanion SJ, Zerbino DR, Abell NS, Akey J, Chen L, Demanelis K, Doherty JA, Feinberg AP, Hansen KD, Hickey PF, Jasmine F, Jiang L, Kaul R, Kibriya MG, Li JB, Li Q, Lin S, Linder SE, Pierce BL, Rizzardi LF, Skol AD, Smith KS, Snyder M, Stamatoyannopoulos J, Tang H, Wang M, Carithers LJ, Guan P, Koester SE, Little AR, Moore HM, Nierras CR, Rao AK, Vaught JB, Volpi S. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science. 2020;369:1318–1330. doi: 10.1126/science.aaz1776. - DOI - PMC - PubMed
-
- Andersson LS, Larhammar M, Memic F, Wootz H, Schwochow D, Rubin C-J, Patra K, Arnason T, Wellbring L, Hjälm G, Imsland F, Petersen JL, McCue ME, Mickelson JR, Cothran G, Ahituv N, Roepstorff L, Mikko S, Vallstedt A, Lindgren G, Andersson L, Kullander K. Mutations in DMRT3 affect locomotion in horses and spinal circuit function in mice. Nature. 2012;488:642–646. doi: 10.1038/nature11399. - DOI - PMC - PubMed
-
- Borgius L, Nishimaru H, Caldeira V, Kunugise Y, Löw P, Reig R, Itohara S, Iwasato T, Kiehn O. Spinal glutamatergic neurons defined by EphA4 signaling are essential components of normal locomotor circuits. The Journal of Neuroscience. 2014;34:3841–3853. doi: 10.1523/JNEUROSCI.4992-13.2014. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 2023YFC2509700/National Key Research and Development Program of China
- 81822030/National Natural Science Foundation of China
- 82102522/National Natural Science Foundation of China
- 31991194 & 32125015/National Natural Science Foundation of China
- 82172382/National Natural Science Foundation of China
- 2021-I2M-1-051/Chinese Academy of Medical Sciences Initiative for Innovative Medicine
- 2021-I2M-1-052/Chinese Academy of Medical Sciences Initiative for Innovative Medicine
- 2020-I2M-C&T-B-030/Chinese Academy of Medical Sciences Initiative for Innovative Medicine
- 7222133/Natural Science Foundation of Beijing Municipality
- No. 2019PT320025/Non-profit Central Research Institute Fund of Chinese Academy of Medical Sciences
- 2022-PUMCH-D-004/National High Level Hospital Clinical Research Funding
- 2022-PUMCH-C-033/National High Level Hospital Clinical Research Funding
- ZR202102210113/Natural Science Foundation of Shandong Province
- X01 HL132375-01A1/Gabriella Miller Kids First Program grant
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
