

This is a preprint.
TDP-43 pathology induces CD8+ T cell activation through cryptic epitope recognition
- PMID: 40667053
- PMCID: PMC12262510
- DOI: 10.1101/2025.06.22.660773
TDP-43 pathology induces CD8+ T cell activation through cryptic epitope recognition
Abstract
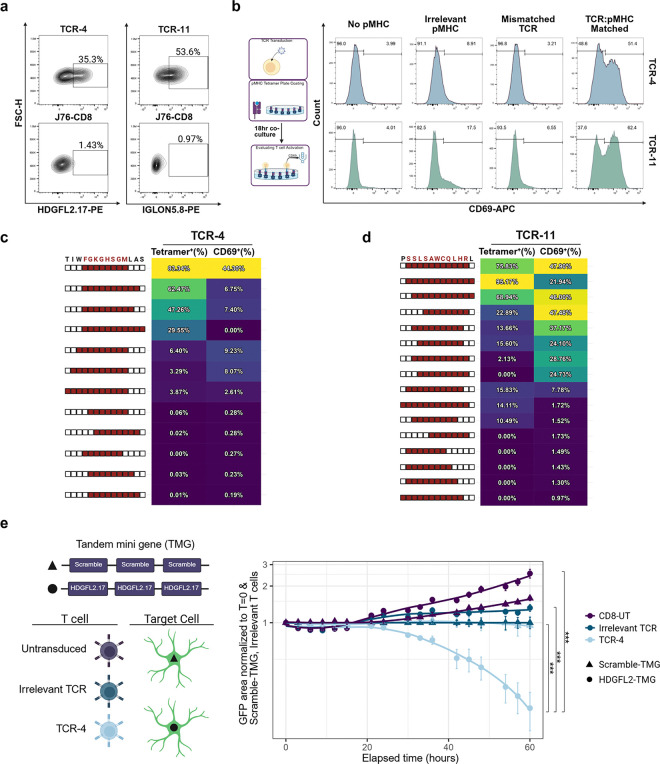
Aggregation and nuclear depletion of the RNA binding protein TDP-43 are the crucial pathological features of amyotrophic lateral sclerosis (ALS) and inclusion body myositis (IBM), two degenerative diseases of the CNS and muscle. The loss of TDP-43 nuclear function results in the aberrant inclusion of cryptic exons in mRNA transcripts, leading to the expression of de novo proteins. Clonally expanded and highly differentiated CD8+ T cells have been observed in individuals with TDP-43 proteinopathies and therapeutics modulating the T cell response have recently been found to extend survival. However, the target antigens mediating T cell activation have remained elusive. Here, we investigate whether the de novo proteins induced by aberrant cryptic splicing due to TDP-43 nuclear loss can act as neo-antigens. We detect the HDGFL2 cryptic peptide and multiple other TDP-43 cryptic exons in IBM skeletal muscle, where their presence correlates with enrichment of T cells and class I antigen presentation pathways. Furthermore, we identify epitopes deriving from HDGFL2 and IGLON5 cryptic peptides which are recognized by clonally expanded and functionally differentiated populations of CD8+ T cells in ALS and IBM Patients. Finally, we demonstrate that T cells engineered to express the identified TCRs can bind and activate in response to the cryptic peptide derived epitopes (cryptic epitopes) and are able to kill TDP-43 deficient astrocytes. This work identifies for the first time specific T cell antigens in ALS and IBM, directly linking adaptive immune response to TDP-43 pathology.
Conflict of interest statement
Competing interests: The University of Pennsylvania has applied for patents on the antigens and TCRs.
Figures





References
-
- Neumann M., Sampathu D. M., Kwong L. K., Truax A. C., Micsenyi M. C., Chou T. T., Bruce J., Schuck T., Grossman M., Clark C. M., McCluskey L. F., Miller B. L., Masliah E., Mackenzie I. R., Feldman H., Feiden W., Kretzschmar H. A., Trojanowski J. Q. & Lee V. M.-Y. Ubiquitinated TDP-43 in Frontotemporal Lobar Degeneration and Amyotrophic Lateral Sclerosis. Science 314, 130–133 (2006). - PubMed
-
- de Boer E. M. J., Orie V. K., Williams T., Baker M. R., De Oliveira H. M., Polvikoski T., Silsby M., Menon P., van den Bos M., Halliday G. M., van den Berg L. H., Van Den Bosch L., van Damme P., Kiernan M., van Es M. A. & Vucic S. TDP-43 proteinopathies: a new wave of neurodegenerative diseases. J. Neurol. Neurosurg. Psychiatry 92, 86–95 (2021). - PMC - PubMed
-
- Brown A. L., Wilkins O. G., Keuss M. J., Hill S. E., Zanovello M., Lee W. C., Bampton A., Lee F. C. Y., Masino L., Qi Y. A., Bryce-Smith S., Gatt A., Hallegger M., Fagegaltier D., Phatnani H., Newcombe J., Gustavsson E. K., Seddighi S., Reyes J. F., Coon S. L., Ramos D., Schiavo G., Fisher E. M. C., Raj T., Secrier M., Lashley T., Ule J., Buratti E., Humphrey J., Ward M. E., Fratta P. & Consortium N. A. TDP-43 loss and ALS-risk SNPs drive mis-splicing and depletion of UNC13A. Nature 603, 131–137 (2022). - PMC - PubMed
-
- Seddighi S., Qi Y. A., Brown A. L., Wilkins O. G., Bereda C., Belair C., Zhang Y. J., Prudencio M., Keuss M. J., Khandeshi A., Pickles S., Kargbo-Hill S. E., Hawrot J., Ramos D. M., Yuan H., Roberts J., Sacramento E. K., Shah S. I., Nalls M. A., Colon-Mercado J. M., Reyes J. F., Ryan V. H., Nelson M. P., Cook C. N., Li Z., Screven L., Kwan J. Y., Mehta P. R., Zanovello M., Hallegger M., Shantaraman A., Ping L., Koike Y., Oskarsson B., Staff N. P., Duong D. M., Ahmed A., Secrier M., Ule J., Jacobson S., Reich D. S., Rohrer J. D., Malaspina A., Dickson D. W., Glass J. D., Ori A., Seyfried N. T., Maragkakis M., Petrucelli L., Fratta P. & Ward M. E. Mis-spliced transcripts generate de novo proteins in TDP-43-related ALS/FTD. Sci Transl Med eadg7162 (2024) doi: 10.1126/scitranslmed.adg7162. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous