

This is a preprint.
The Luminal Ring Protein C2CD3 Acts as a Radial In-to-Out Organizer of the Distal Centriole and Appendages
- PMID: 40667239
- PMCID: PMC12262607
- DOI: 10.1101/2025.06.17.660204
The Luminal Ring Protein C2CD3 Acts as a Radial In-to-Out Organizer of the Distal Centriole and Appendages
Update in
-
The luminal ring protein C2CD3 acts as a radial in-to-out organizer of the distal centriole and appendages.PLoS Biol. 2025 Dec 9;23(12):e3003519. doi: 10.1371/journal.pbio.3003519. eCollection 2025 Dec. PLoS Biol. 2025. PMID: 41364719 Free PMC article.
Abstract
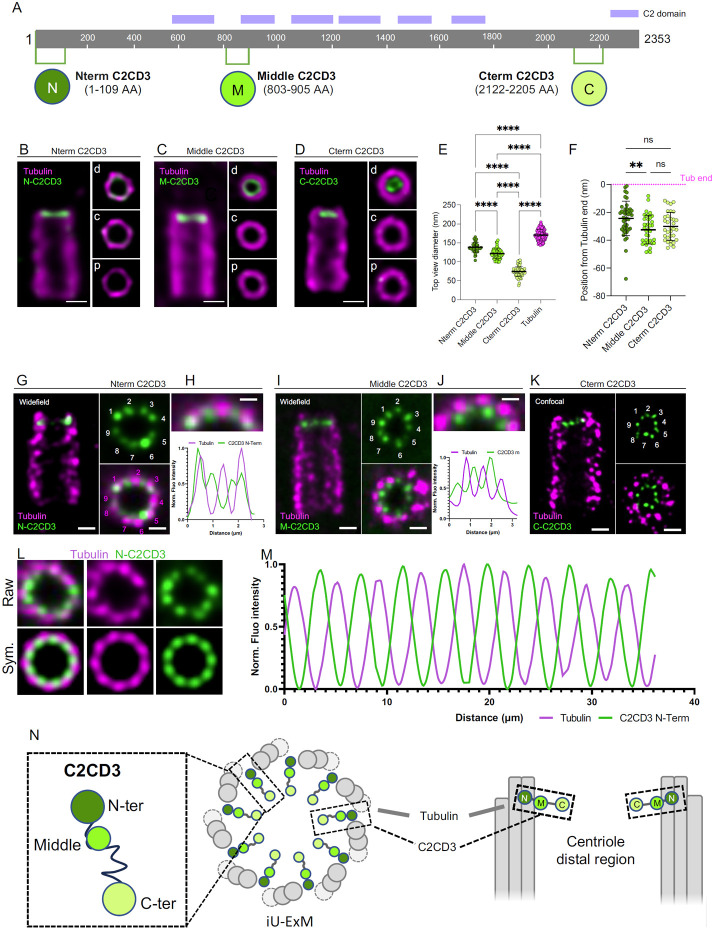
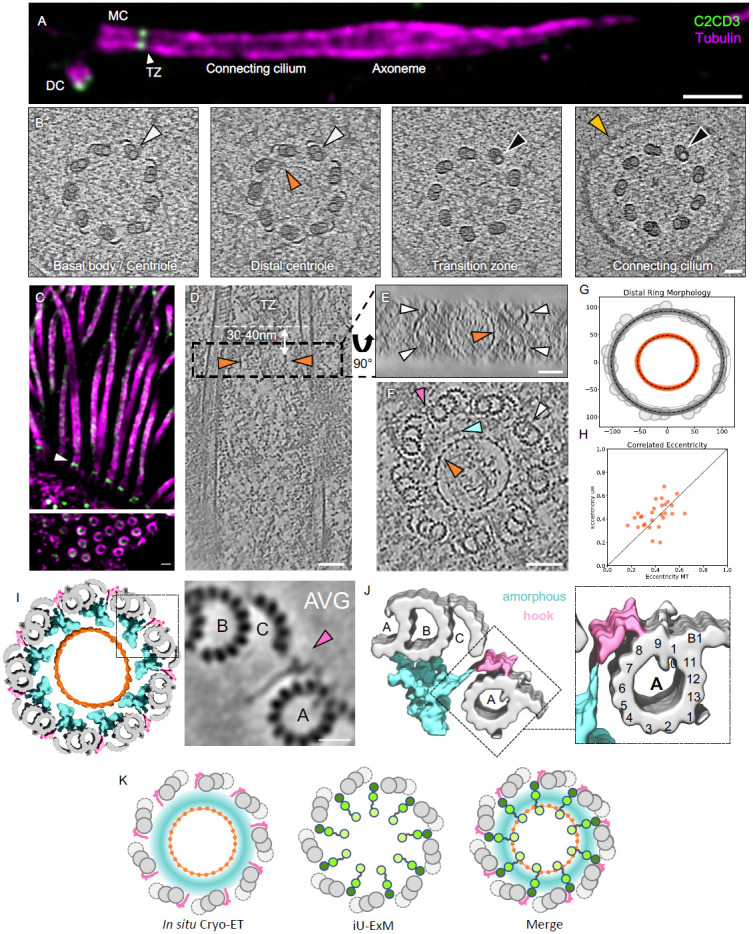
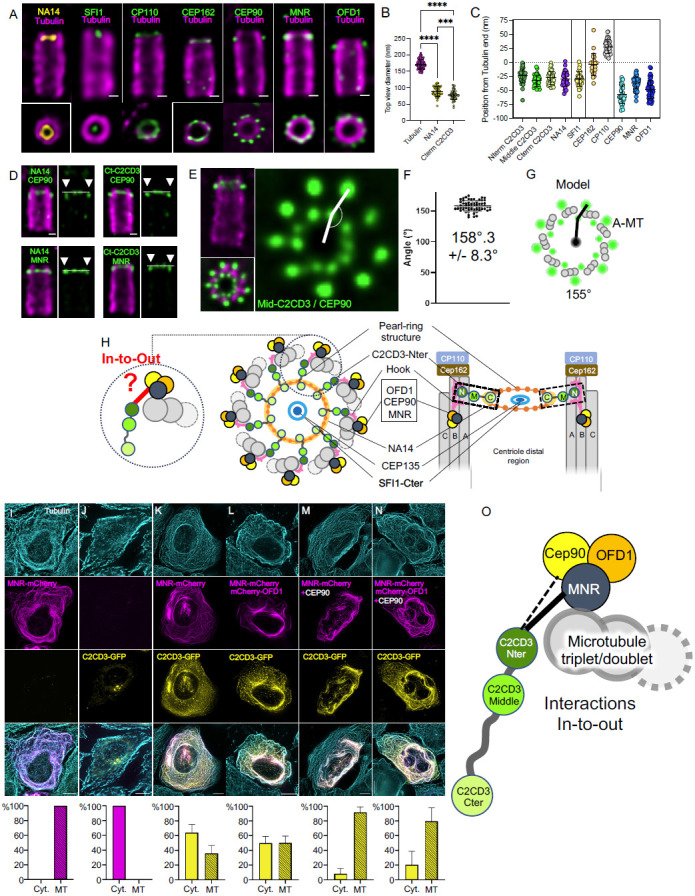
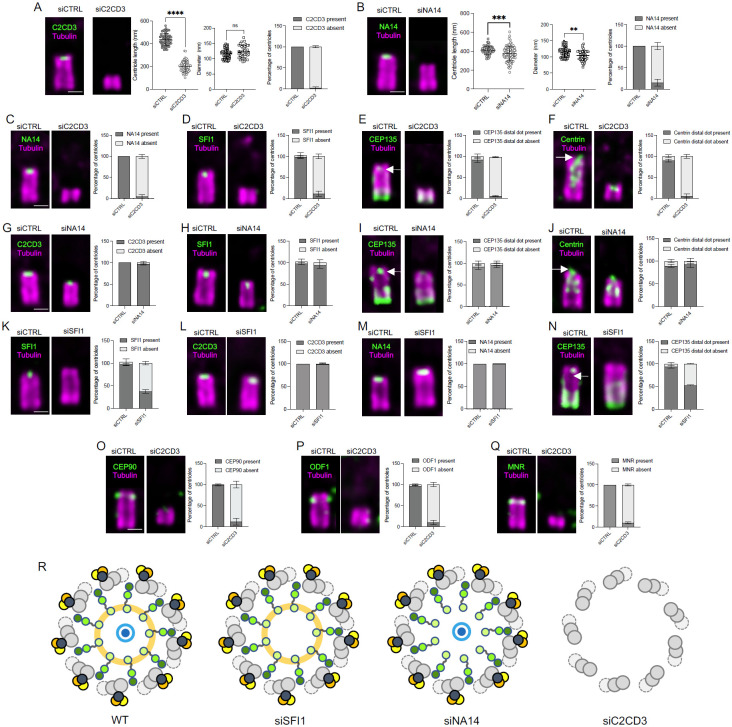
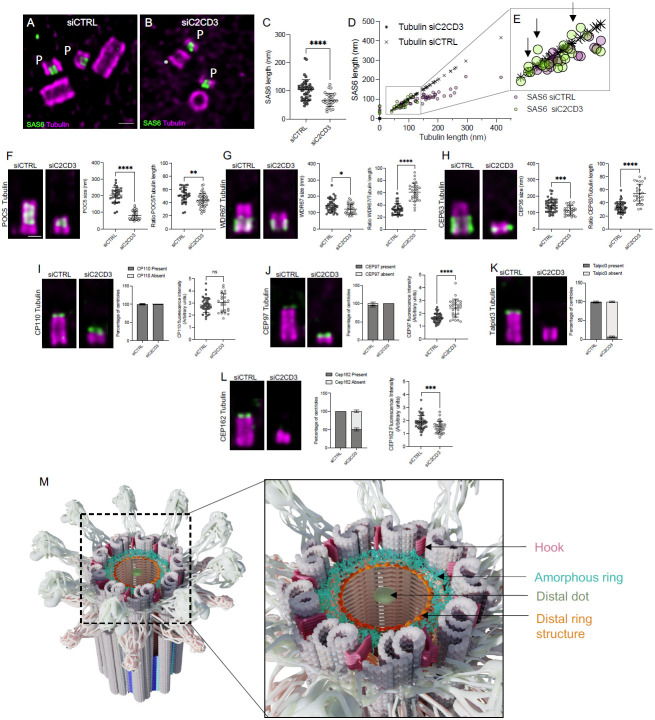
Centrioles are polarized microtubule-based structures with appendages at their distal end that are essential for cilia formation and function. The protein C2CD3 is critical for distal appendage assembly, with mutations linked to orofaciodigital syndrome and other ciliopathies. However, its precise molecular role in appendage recruitment remains unclear. Using Ultrastructure Expansion Microscopy (U-ExM), iterative U-ExM, and in situ cryo-electron tomography (cryo-ET), we reveal that C2CD3 adopts a radially symmetric 9-fold organization within the centriole's distal lumen. We show that the C-terminal region of C2CD3 localizes close to a ~100 nm luminal ring structure consisting of ~27 nodes, while its N-terminal region localizes close to a hook-like structure that attaches to the A-microtubule as it extends from the centriole interior to exterior. This hook structure is adjacent to the DISCO complex (MNR/CEP90/OFD1), which marks future appendage sites. C2CD3 depletion disrupts not only the recruitment of the DISCO complex via direct interaction with MNR but also destabilizes the luminal ring network composed of C2CD3/SFI1/centrin-2/CEP135/NA14, as well as the distal microtubule tip protein CEP162. This reveals an intricate "in-to-out" molecular hub connecting the centriolar lumen, distal microtubule cap, and appendages. Although C2CD3 loss results in shorter centrioles and appendage defects, key structural elements remain intact, permitting continued centriole duplication. We propose that C2CD3 forms the luminal ring structure and extends radially to the space between triplet microtubules, functioning as an architectural hub that scaffolds the distal end of the centriole, orchestrating its assembly and directing appendage formation.
Keywords: C2CD3; appendages; centriole; expansion microscopy; in situ cryo-ET; luminal distal ring.
Conflict of interest statement
Conflict of interest The authors declare no competing interests.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous