

Single-cell spatial analysis with Xenium reveals anti-tumour responses of CXCL13 + T and CXCL9+ cells after radiotherapy combined with anti-PD-L1 therapy
- PMID: 40670667
- PMCID: PMC12449477
- DOI: 10.1038/s41416-025-03088-0
Single-cell spatial analysis with Xenium reveals anti-tumour responses of CXCL13 + T and CXCL9+ cells after radiotherapy combined with anti-PD-L1 therapy
Abstract
Background: The standard treatment for unresectable non-small cell lung cancer (NSCLC) is anti-PD-L1 therapy combined with chemoradiotherapy (anti-PD-L1-CRT). Although some patients achieve complete cancer eradication and cure, more than half of patients retain persistent cancer cells. Our research aimed to unravel the nuanced mechanisms involved in both immune attack and evasion induced by anti-PD-L1-CRT with single cell spatial transcriptome.
Methods: Xenium is a cutting-edge single-cell spatial analysis tool that enables pathology-based and single-cell analyses while preserving spatial information. In our study, we used Xenium to identify the tumour microenvironment (TME), immune dynamics, and residual cancer cells at the single-cell level following treatment with anti-PD-L1-CRT.
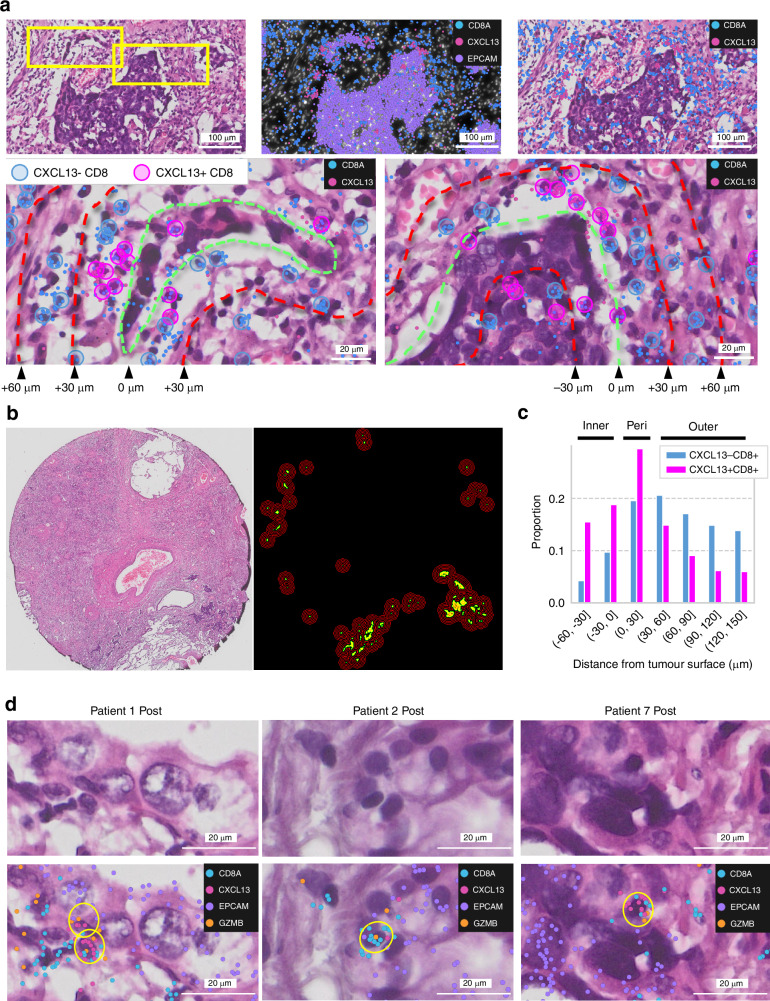
Results: Posttreatment alterations included a significant increase in CXCL9+ cells and CXCL13 + T cells, particularly around tumour cells. Additionally, we discovered that CXCL13 + T cells directly impact cancer cells in the posttreatment environment. Moreover, we identified clusters of immune-cold cancer cells posttreatment, revealing their activation of DNA repair pathways and high proliferative capacity. The novel spatial analysis tool Xenium enabled identification of the immune environment at the single-cell level following treatment with anti-PD-L1-CRT, elucidating its characteristics.
Conclusions: These findings suggest potential advancements in developing new treatments to improve posttreatment immune responses and address resistance challenges.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: M.N. reports personal fees from AstraZeneca and a research grant from illumina, outside the submitted work. T.K. received research grant from Roche Diagnostics K.K. outside of this work. M.T. has received honorarium and lecture fees from Johnson & Johnson Japan, AstraZeneca KK, Eli Lilly Japan, Chugai Pharmaceutical CO., LTD, Taiho Pharma, Medtronic Japan, Ono Pharmaceutical CO., LTD, MSD, Bristol-Myers Squibb KK, Novartis, Amgen KK, and Daiichi-Sankyo. Additionally, M.T. has been involved in consulting or advisory roles with AstraZeneca KK, Chugai Pharmaceutical CO., LTD, MSD, and Novartis, and has received research grants and engaged in commissioned research (e.g., clinical trials) from Boehringer-Ingelheim Japan, MSD, AstraZeneca KK, Ono Pharmaceutical CO., LTD, MSD, Bristol-Myers Squibb KK, Novartis, Eli Lilly Japan, and MiRXES. T.M. has received honorarium and lecture fees from AstraZeneca KK, Chugai Pharmaceutical CO., LTD, Ono Pharmaceutical CO., LTD, and Bristol-Myers Squibb KK, MSD, Boehringer Ingelheim, Taiho, Eli-Lilly, Novartis, Amgen, Takeda, Kyorin and receieved research grant from AstraZeneca, Chugai, Boeringer Ingelheim, Pfizer, Taiho, MSD, Ono, Bridge Biopharma and Natera. A.H. has received honorarium and lecture fees from AstraZeneca KK, Chugai Pharmaceutical CO., LTD, Ono Pharmaceutical CO., LTD, and Bristol-Myers Squibb KK. Additionally, A.H. has been involved in consulting or advisory roles with AstraZeneca KK and has received research grants and engaged in commissioned research (e.g., clinical trials) from AstraZeneca KK. J.S. received honorarium and lecture fees from Johnson & Johnson Japan, Chugai Pharmaceutical CO., LTD, Taiho Pharma, Medtronic Japan, and Intuitive Japan. Ethics approval and consent to participate: Ethical approval was obtained from the ethics committee of National Cancer Center Japan (Protocol Number: 2022-407). The study was conducted in accordance with the principles of the Declaration of Helsinki.
Figures





References
-
- Galluzzi L, Buqué A, Kepp O, Zitvogel L, Kroemer G. Immunogenic cell death in cancer and infectious disease. Nat Rev Immunol. 2017;17:97–111. - PubMed
-
- Bando H, Tsukada Y, Ito M, Yoshino T. Novel immunological approaches in the treatment of locally advanced rectal cancer. Clin Colorectal Cancer. 2022;21:3–9. - PubMed
-
- Zhu X, Cao Y, Liu W, Ju X, Zhao X, Jiang L, et al. Stereotactic body radiotherapy plus pembrolizumab and trametinib versus stereotactic body radiotherapy plus gemcitabine for locally recurrent pancreatic cancer after surgical resection: an open-label, randomised, controlled, phase 2 trial. Lancet Oncol. 2021;22:1093–102. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
