

Identification of BAHD-acyltransferase enzymes involved in ingenane diterpenoid biosynthesis
- PMID: 40671640
- PMCID: PMC12371174
- DOI: 10.1111/nph.70388
Identification of BAHD-acyltransferase enzymes involved in ingenane diterpenoid biosynthesis
Abstract
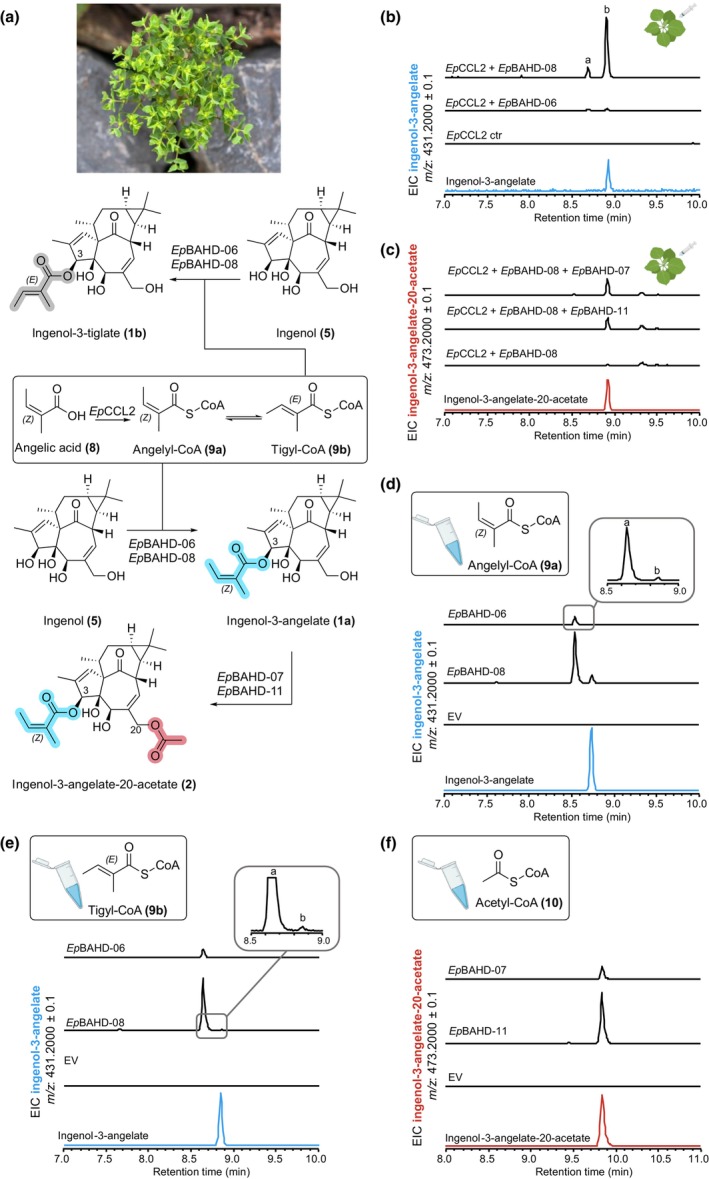
The plant family Euphorbiaceae is an abundant source of structurally complex diterpenoids, many of which have reported anticancer, anti-HIV, and anti-inflammatory activities. Among these, ingenol-3-angelate (1a; tradename: Picato®), isolated from Euphorbia peplus, has potent antitumor activity. We report the discovery and characterization of the first genes linked to committed steps of ingenol-3-angelate (1a) biosynthesis in E. peplus. Using pathway reconstitution in Nicotiana benthamiana and in vitro assays with recombinant enzymes, we identified two genes whose products catalyze the addition of angelyl-CoA (9a) to the ingenol (5) scaffold, producing ingenol-3-angelate (1a). We also identified three genes whose products catalyze acetylation of ingenol-3-angelate (1a) to ingenol-3-angelate-20-acetate (2). Virus induced gene silencing (VIGS) suggests considerable functional redundancy in the E. peplus genome for this enzymatic step. We also identified three genes whose products can catalyze acetylation of ingenol-3-angelate (1a) to ingenol-3-angelate-20-acetate (2). In this case, virus-induced gene silencing (VIGS) indicates considerable functional redundancy in the E. peplus genome of genes encoding this enzymatic step. We demonstrate using VIGS that just one of these genes, EpBAHD-08, is essential for this angeloylation in E. peplus. VIGS of the second gene, EpBAHD-06, has a significant effect on jatrophanes rather than ingenanes in E. peplus. This work paves the way for increasing ingenol-3-angelate (1a) levels in planta and provides a foundation for the discovery of the remaining genes in the biosynthetic pathway of these important molecules.
Keywords: BAHD‐acyltransferase; Euphorbia peplus; diterpene; ingenol‐3‐angelate; medicinal plant; natural product biosynthesis.
© 2025 The Author(s). New Phytologist © 2025 New Phytologist Foundation.
Conflict of interest statement
None declared.
Figures



Similar articles
-
Dietary cancer risk from conditional cancerogens in produce of livestock fed on species of spurge (Euphorbiaceae). III. Milk of lactating goats fed on the skin irritant herb Euphorbia peplus is polluted by tumor promoters of the ingenane diterpene ester type.J Cancer Res Clin Oncol. 1998;124(6):301-6. doi: 10.1007/s004320050173. J Cancer Res Clin Oncol. 1998. PMID: 9692836 Free PMC article.
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Two O-acyltransferases from the diterpene biosynthetic gene cluster of Euphorbia lathyris contribute to the structural diversity of medicinal macrocyclic diterpenoid esters biosynthesis.Plant J. 2025 Feb;121(4):e70003. doi: 10.1111/tpj.70003. Plant J. 2025. PMID: 39968625
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2020 Jan 9;1(1):CD011535. doi: 10.1002/14651858.CD011535.pub3. Cochrane Database Syst Rev. 2020. Update in: Cochrane Database Syst Rev. 2021 Apr 19;4:CD011535. doi: 10.1002/14651858.CD011535.pub4. PMID: 31917873 Free PMC article. Updated.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
References
-
- Alves ALV, Da Silva LS, Faleiros CA, Silva VAO, Reis RM. 2022. The role of ingenane diterpenes in cancer therapy: from bioactive secondary compounds to small molecules. Natural Product Communications 17: 1934578X2211056.
-
- Czechowski T, Forestier E, Swamidatta SH, Gilday AD, Cording A, Larson TR, Harvey D, Li Y, He Z, King AJ et al. 2022. Gene discovery and virus‐induced gene silencing reveal branched pathways to major classes of bioactive diterpenoids in Euphorbia peplus . Proceedings of the National Academy of Sciences, USA 119: e2203890119. - PMC - PubMed
-
- D'Auria JC. 2006. Acyltransferases in plants: a good time to be BAHD. Current Opinion in Plant Biology 9: 331–340. - PubMed
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
