

This is a preprint.
Lysosomal Protease-Mediated APP Degradation is pH-Dependent, Mutation-Sensitive, and Facilitates Tau Proteolysis
- PMID: 40671818
- PMCID: PMC12265180
- DOI: 10.21203/rs.3.rs-6978813/v1
Lysosomal Protease-Mediated APP Degradation is pH-Dependent, Mutation-Sensitive, and Facilitates Tau Proteolysis
Abstract
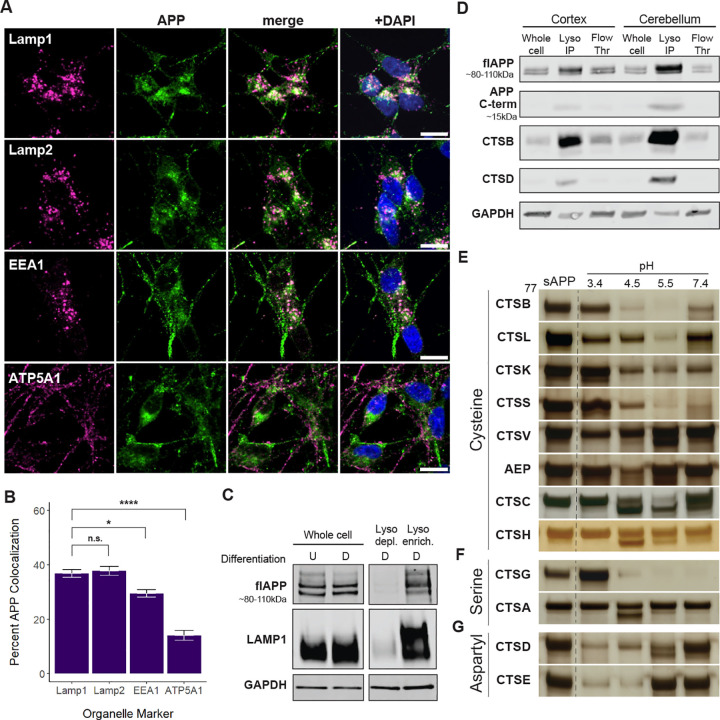
Background: The accumulation and aggregation of amyloid beta (Aβ)-a peptide fragment derived from the proteolytic processing of amyloid precursor protein (APP)-is a central pathological feature of Alzheimer's disease (AD) and a current target for disease-modifying therapies. Mutations in APP can also drive early-onset AD. While the roles of α-, β-, and γ-secretases and their respective cleavage sites in APP processing are well characterized, much less is understood about the routine degradation of APP within sub-cellular compartments like the lysosome.
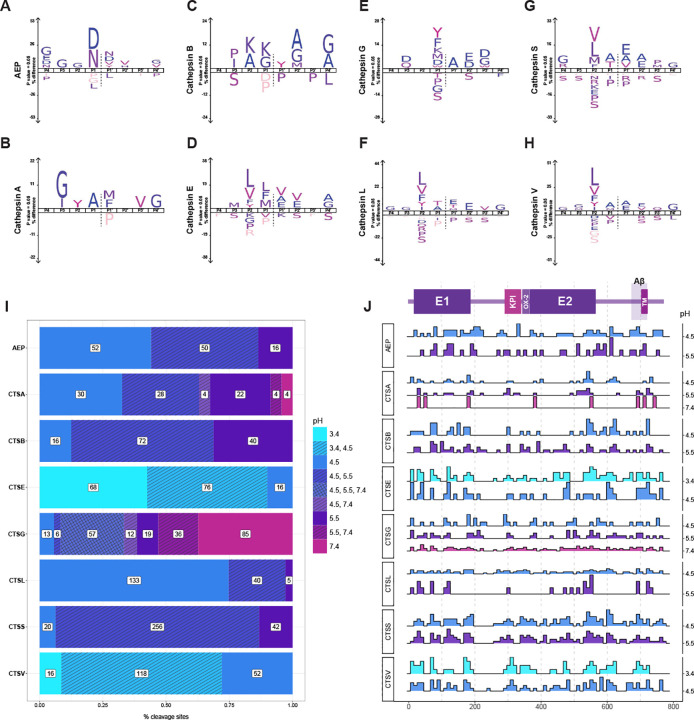
Methods: We applied Multiplexed Substrate Profiling by Mass Spectrometry (MSP-MS) to map cleavage sites within APP that may be targeted by lysosomal proteases, also known as cathepsins. We then employed cell-based and in vitro assays to examine the degradation of both wild-type and mutant APP by these enzymes.
Results: Our findings confirm that APP is enriched in the endolysosomal compartment, where it is processed by many, but not all, cathepsins. Our experiments reveal that cleavages at several mapped APP sites are sensitive to both changes in pH and the presence of pathogenic variants E693G and E693Q. Additionally, we discovered that the large soluble domain of APP (sAPP) enhances tau cleavage by cathepsin G in vitro.
Conclusions: Collectively, these results underscore the importance of lysosomal processing of APP, identify a link between APP and tau, and suggest new avenues for exploring AD pathogenesis. They also point to potential therapeutic targets related to the lysosomal function of APP and its impact on neurodegenerative disease.
Keywords: APP; Alzheimer’s disease; Aβ; amyloid-beta; amyloid-precursor protein; autophagy; cathepsin; lysosome; neurodegeneration; protease; tau.
Conflict of interest statement
Competing interests A.W.K. is a Scientific Advisory Board members, and G.M.K. is an employee of Alaunus Biosciences. Authors declare that they have no conflicts of interest or competing interests.
Figures



References
-
- Kimura A, Hata S, Suzuki T. Alternative Selection of β-Site APP-Cleaving Enzyme 1 (BACE1) Cleavage Sites in Amyloid β-Protein Precursor (APP) Harboring Protective and Pathogenic Mutations within the Aβ Sequence*. Journal of Biological Chemistry. 2016. Nov;291(46):24041–24053. 10.1074/jbc.M116.744722. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
