

This is a preprint.
It has not yet been peer reviewed by a journal.
The National Library of Medicine is
running a pilot
to include preprints that result from research funded by NIH in PMC and PubMed.
[Preprint]. 2025 Jul 11:2025.07.11.664319.
doi: 10.1101/2025.07.11.664319.
c-di-GMP-Dependent Regulation of Motility by comFB and comFC
Affiliations
- PMID: 40672238
- PMCID: PMC12265714
- DOI: 10.1101/2025.07.11.664319
Item in Clipboard
c-di-GMP-Dependent Regulation of Motility by comFB and comFC
bioRxiv.
.
Abstract
ComFB is encoded in the comF operon of Bacillus subtilis, situated between the genes for ComFA and ComFC. The latter two proteins are essential for natural transformation, whereas ComFB is dispensable. We show here that ComFB binds specifically and with high affinity to the second messenger c-di-GMP and that ComFB acts as a c-di-GMP receptor to inhibit swarming and swimming motility, apparently by interfering with flagellar activity. We show further that in the absence of ComFC, swarming is completely abrogated by a mechanism that requires FB. These results reveal a new c-di-GMP regulatory system that controls motility independently of MotI.
Figures
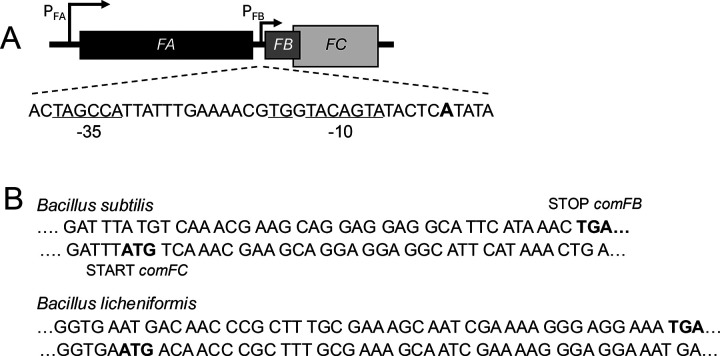
Organization of the comF operon. (A) In Bsu, the comFA, comFB, and comFC open reading frames are transcribed from PFA, a ComK-dependent promoter. PFB, a ComK-independent promoter, drives comFB and FC, which overlap. The existence of PFB is documented as discussed in the text. The +1 A-residue for PFB is shown in boldface, and the proposed promoter sequence features are underlined. (B) Two examples of bacteria that encode ComFB in their comF operons, demonstrating the overlap of their comFB and comFC coding sequences.
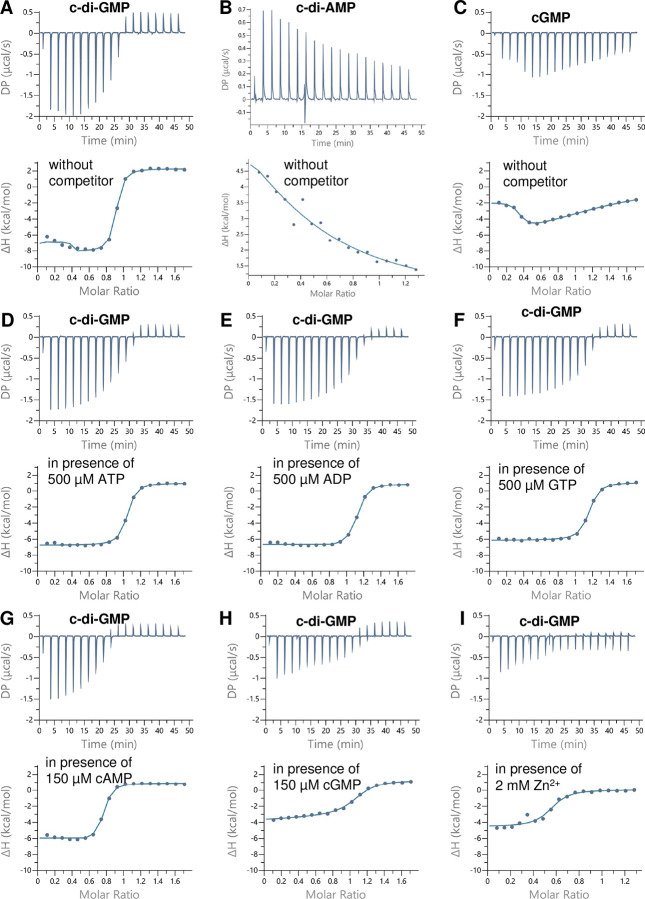
Figure 2. Isothermal titration calorimetry (ITC) analysis of c-di-GMP, cGMP, and c-di-AMP binding to FB, as indicated. Upper panels show the raw ITC data in the form of heat produced during the titration of the respective nucleotide on the FB protein; lower panels show the binding isotherms and the best-fit curves according to the one binding site model. (A-C) ITC analysis of c-di-GMP (A), c-di-AMP (B), and cGMP (C) binding to FB in the absence of competitors. (D-I) ITC analysis of c-di-GMP binding to FB in the presence of ATP (D), ADP (E), GTP (F), cAMP (G), cGMP (H), or Zn2+ (I).

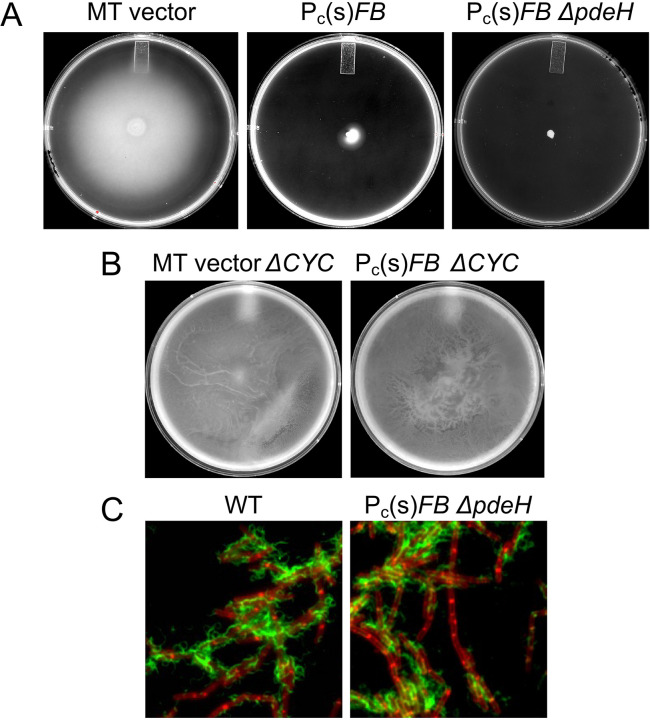
FB, together with c-di-GMP, inhibits swarming. Each of the panels is a representative of at least three repeated swarming experiments, performed as described using 0.7% LB-agar plates. The expansion diameter of the cell sheet was measured at intervals and plotted against time. The symbols are consistent across all five panels, with empty symbols indicating ΔpdeH strains. MT: empty vector (pKB149 (33)) in amyE. The constructs amyE::Pc(w)FB and amyE::Pc(s)FB are described in the text. Δcyc denotes strains deleted for the three diguanlylate cyclases of Bsu (dgcP, dgcK, dgcW).

The expression of FB and its regulation. Western blots were performed with an anti-serum raised against purified FB. As a loading control, the tops of the membranes were cut off after blotting and developed with an antiserum raised against elongation factor G (EFG). We have noticed a consistent signal, indicated by arrows, at the position expected for a dimer of FB. The Δ(FB-FC) strain was included in each panel to confirm the identity of the FB signal. The vertical dashed lines indicate that intervening lanes were omitted from a gel. For the gel to the right of panel A, the first three lanes were loaded with lysates diluted 20-fold, and only 2 μl were loaded compared to 10–20 μl of undiluted lysate for the other lanes. This is evident from the weaker EFG signals. The empty vector strain produces a low level of FB, which is increased moderately in the amyE::Pc(w)FB strain and about 100-fold over the wild-type level in the amyE::Pc(s)FB strain.
FB inhibits swimming but not the production of flagella. (A and B) The indicated strains were tested for swimming in 0.3% agar. The images were collected when the empty vector (MT) control strains had reached or nearly reached the outer limits of the Petri dishes. (C) The two indicated strains, both carrying the hagT209C allele, were reacted with Alexa Fluor 488 C5 maleimide (green), stained with the membrane dye FM4–64 (red), and imaged as described in Materials and Methods.
Similar articles
-
ComFB, a new widespread family of c-di-NMP receptor proteins.bioRxiv [Preprint]. 2024 Nov 10:2024.11.10.622515. doi: 10.1101/2024.11.10.622515. bioRxiv. 2024. PMID: 39574629 Free PMC article. Preprint.
-
The cyclic di-GMP receptor YcgR links the second messenger with the putrescine quorum sensing system in modulation of Dickeya oryzae motility.mBio. 2025 Jul 9;16(7):e0101625. doi: 10.1128/mbio.01016-25. Epub 2025 May 30. mBio. 2025. PMID: 40444987 Free PMC article.
-
Functional analysis of cyclic diguanylate-modulating proteins in Vibrio fischeri.mSystems. 2024 Nov 19;9(11):e0095624. doi: 10.1128/msystems.00956-24. Epub 2024 Oct 22. mSystems. 2024. PMID: 39436151 Free PMC article.
-
Antioxidants for male subfertility.Cochrane Database Syst Rev. 2014;(12):CD007411. doi: 10.1002/14651858.CD007411.pub3. Epub 2014 Dec 15. Cochrane Database Syst Rev. 2014. Update in: Cochrane Database Syst Rev. 2019 Mar 14;3:CD007411. doi: 10.1002/14651858.CD007411.pub4. PMID: 25504418 Updated.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
References
-
- Jenal U, Reinders A, Lori C. 2017. Cyclic di-GMP: second messenger extraordinaire. Nat Rev Microbiol 15:271–284. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources