

Structures of Listeria monocytogenes MenD in ThDP-bound and in-crystallo captured intermediate I-bound forms
- PMID: 40673487
- PMCID: PMC12312563
- DOI: 10.1107/S2053230X25006181
Structures of Listeria monocytogenes MenD in ThDP-bound and in-crystallo captured intermediate I-bound forms
Abstract
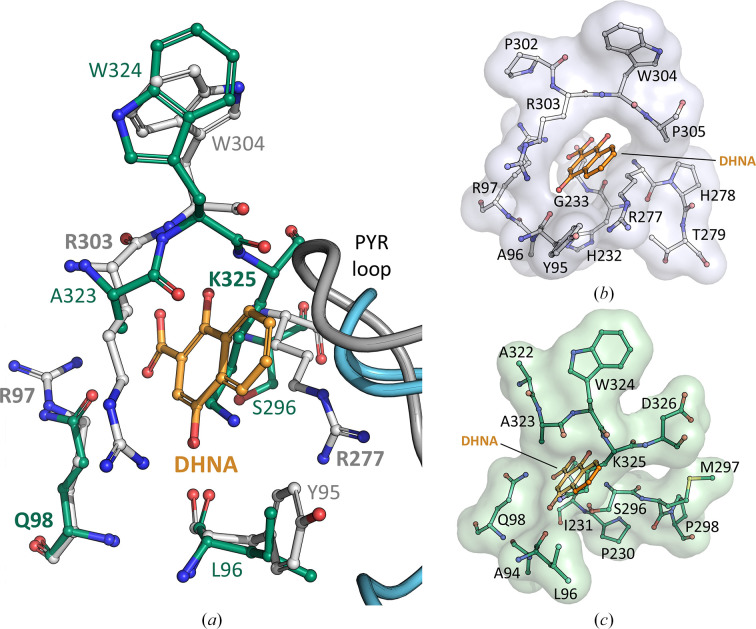
Menaquinones (vitamin K2) are a family of redox-active small lipophilic molecules that serve as vital electron carriers in many bacterial electron-transport pathways. The ThDP-dependent enzyme 2-succinyl-5-enolpyruvyl-6-hydroxy-3-cyclohexene-1-carboxylate (SEPHCHC) synthase (MenD) catalyses the first irreversible step in bacterial classical menaquinone biosynthesis via a series of reactions involving covalent ThDP-bound intermediates. We report structures of MenD from the pathogen Listeria monocytogenes (LmoMenD) in its ThDP cofactor-bound and in-crystallo captured intermediate I-bound forms. Analysis of the structures revealed that LmoMenD adopts the typical three-domain ThDP-dependent fold observed for MenD orthologs, while a combination of structure, size-exclusion chromatography, mass photometry and small-angle X-ray scattering analysis showed that the enzyme has a homotetrameric quaternary structure. While both of the ligand-bound structures reported here were very similar, comparison with an apo form from the PDB revealed a closing down of the active site in the ligand-bound forms, with more complete models suggesting lower levels of disorder around key regions of the active site that interface with ThDP or the captured intermediate. Enzyme kinetics characterization showed the enzyme was active and enabled allosteric inhibition to be measured. There was weak inhibition of enzyme activity in the presence of 1,4-dihydroxy-2-naphthoic acid, an allosteric regulator of Mycobacterium tuberculosis MenD and downstream metabolite in the menaquinone-biosynthesis pathway.
Keywords: Listeria monocytogenes; MenD; SEPHCHC synthase; intermediate I capture; menaquinone biosynthesis; thiamine diphosphate-dependent enzyme.
open access.
Figures



Similar articles
-
Allosteric regulation of menaquinone (vitamin K2) biosynthesis in the human pathogen Mycobacterium tuberculosis.J Biol Chem. 2020 Mar 20;295(12):3759-3770. doi: 10.1074/jbc.RA119.012158. Epub 2020 Feb 6. J Biol Chem. 2020. PMID: 32029475 Free PMC article.
-
Specificity and reactivity in menaquinone biosynthesis: the structure of Escherichia coli MenD (2-succinyl-5-enolpyruvyl-6-hydroxy-3-cyclohexadiene-1-carboxylate synthase).J Mol Biol. 2008 Dec 31;384(5):1353-68. doi: 10.1016/j.jmb.2008.10.048. Epub 2008 Nov 1. J Mol Biol. 2008. PMID: 18983854 Free PMC article.
-
Two active site arginines are critical determinants of substrate binding and catalysis in MenD: a thiamine-dependent enzyme in menaquinone biosynthesis.Biochem J. 2018 Nov 30;475(22):3651-3667. doi: 10.1042/BCJ20180548. Biochem J. 2018. PMID: 30341164
-
Impact of residual disease as a prognostic factor for survival in women with advanced epithelial ovarian cancer after primary surgery.Cochrane Database Syst Rev. 2022 Sep 26;9(9):CD015048. doi: 10.1002/14651858.CD015048.pub2. Cochrane Database Syst Rev. 2022. PMID: 36161421 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
References
-
- Agirre, J., Atanasova, M., Bagdonas, H., Ballard, C. B., Baslé, A., Beilsten-Edmands, J., Borges, R. J., Brown, D. G., Burgos-Mármol, J. J., Berrisford, J. M., Bond, P. S., Caballero, I., Catapano, L., Chojnowski, G., Cook, A. G., Cowtan, K. D., Croll, T. I., Debreczeni, J. É., Devenish, N. E., Dodson, E. J., Drevon, T. R., Emsley, P., Evans, G., Evans, P. R., Fando, M., Foadi, J., Fuentes-Montero, L., Garman, E. F., Gerstel, M., Gildea, R. J., Hatti, K., Hekkelman, M. L., Heuser, P., Hoh, S. W., Hough, M. A., Jenkins, H. T., Jiménez, E., Joosten, R. P., Keegan, R. M., Keep, N., Krissinel, E. B., Kolenko, P., Kovalevskiy, O., Lamzin, V. S., Lawson, D. M., Lebedev, A. A., Leslie, A. G. W., Lohkamp, B., Long, F., Malý, M., McCoy, A. J., McNicholas, S. J., Medina, A., Millán, C., Murray, J. W., Murshudov, G. N., Nicholls, R. A., Noble, M. E. M., Oeffner, R., Pannu, N. S., Parkhurst, J. M., Pearce, N., Pereira, J., Perrakis, A., Powell, H. R., Read, R. J., Rigden, D. J., Rochira, W., Sammito, M., Sánchez Rodríguez, F., Sheldrick, G. M., Shelley, K. L., Simkovic, F., Simpkin, A. J., Skubak, P., Sobolev, E., Steiner, R. A., Stevenson, K., Tews, I., Thomas, J. M. H., Thorn, A., Valls, J. T., Uski, V., Usón, I., Vagin, A., Velankar, S., Vollmar, M., Walden, H., Waterman, D., Wilson, K. S., Winn, M. D., Winter, G., Wojdyr, M. & Yamashita, K. (2023). Acta Cryst. D79, 449–461.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
