

Teasing apart the sources of phylogenetic tree discordance across three genomes in the oak family (Fagaceae)
- PMID: 40676558
- PMCID: PMC12269116
- DOI: 10.1186/s12870-025-06963-3
Teasing apart the sources of phylogenetic tree discordance across three genomes in the oak family (Fagaceae)
Abstract
Background: Gene tree incongruence is a well-documented, but the biological and analytical factors driving phylogenetic discordance remains incompletely understood. In this study, we investigated how different factors contribute to incongruence among gene trees in Fagaceae.
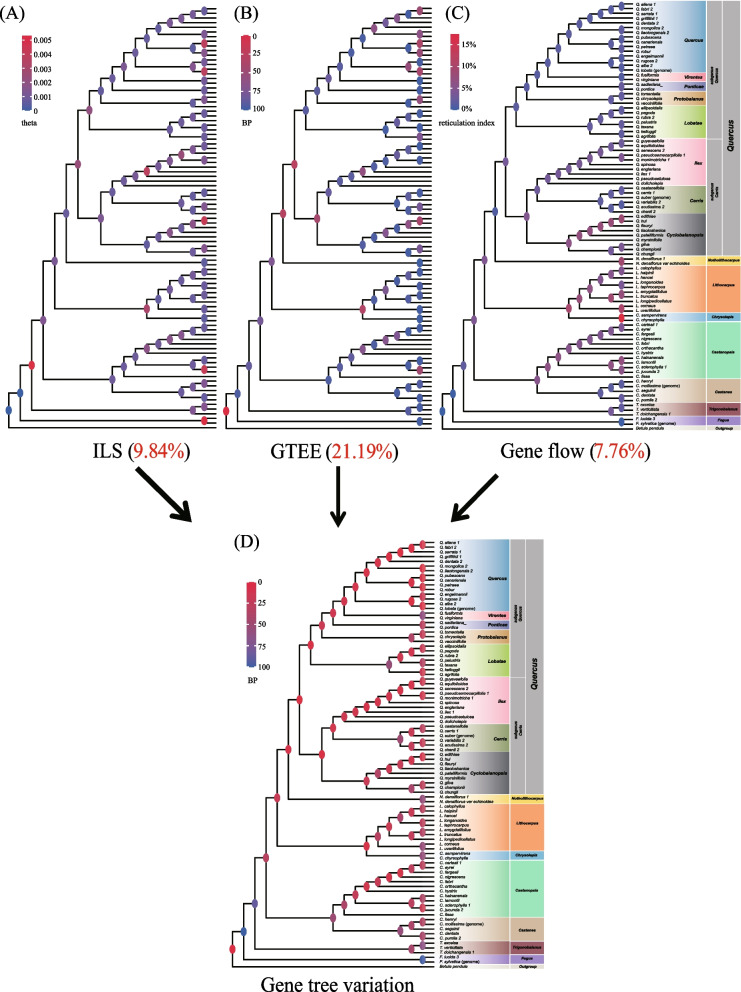
Results: Each dataset produced highly supported topologies, with Fagus and Trigonobalanus consistently placed as early-diverging lineages within the Fagaceae family. However, the cpDNA and mtDNA divided the remaining Fagaceae species into New World and Old World clades, a pattern that sharply contrasted with the phylogenetic relationships inferred from nuclear genome data. These discrepancies between the cytoplasmic and nuclear gene trees likely result from ancient interspecific hybridization within Fagaceae. The decomposition analyses revealed that gene tree estimation error, incomplete lineage sorting, and gene flow accounted for 21.19%, 9.84%, and 7.76% of gene tree variation, respectively. We further revealed that 58.1-59.5% of genes exhibited consistent phylogenetic signals ("consistent genes"), while 40.5-41.9% of genes displayed conflicting signals ("inconsistent genes"). Consistent genes showed stronger phylogenetic signals and were more likely to recover the species tree topology than inconsistent genes. However, consistent and inconsistent genes did not significantly differ in terms of sequence- and tree-based characteristics. By excluding a subset of inconsistent genes, the study significantly reduced inconsistencies between concatenation- and coalescent-based approaches.
Conclusions: This study illustrates how diverse factors contribute to gene tree incongruence, offering new insights into the evolutionary history of Fagaceae.
Keywords: Fagaceae; Gene tree discordance; Gene tree estimation error; Hybridization; Incomplete lineage sorting.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: The collection of all samples fully complied with national and local legislation. In accordance with national and local laws and regulations, special permission was granted for the collection of plant samples in this study. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures




References
-
- Kapli P, Yang Z, Telford MJ. Phylogenetic tree building in the genomic age. Nat Rev Genet. 2020;21(7):428–44. - PubMed
-
- Shao Y, Zhou L, Li F, Zhao L, Zhang BL, Shao F, Chen JW, Chen CY, Bi XP, Zhuang XL, et al. Phylogenomic analyses provide insights into primate evolution. Science. 2023;380(6648):913–24. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
