

Synergistic activation of bat SARS-like coronaviruses spike protein by elastase and TMPRSS2
- PMID: 40691229
- PMCID: PMC12279935
- DOI: 10.1038/s41598-025-11600-y
Synergistic activation of bat SARS-like coronaviruses spike protein by elastase and TMPRSS2
Abstract
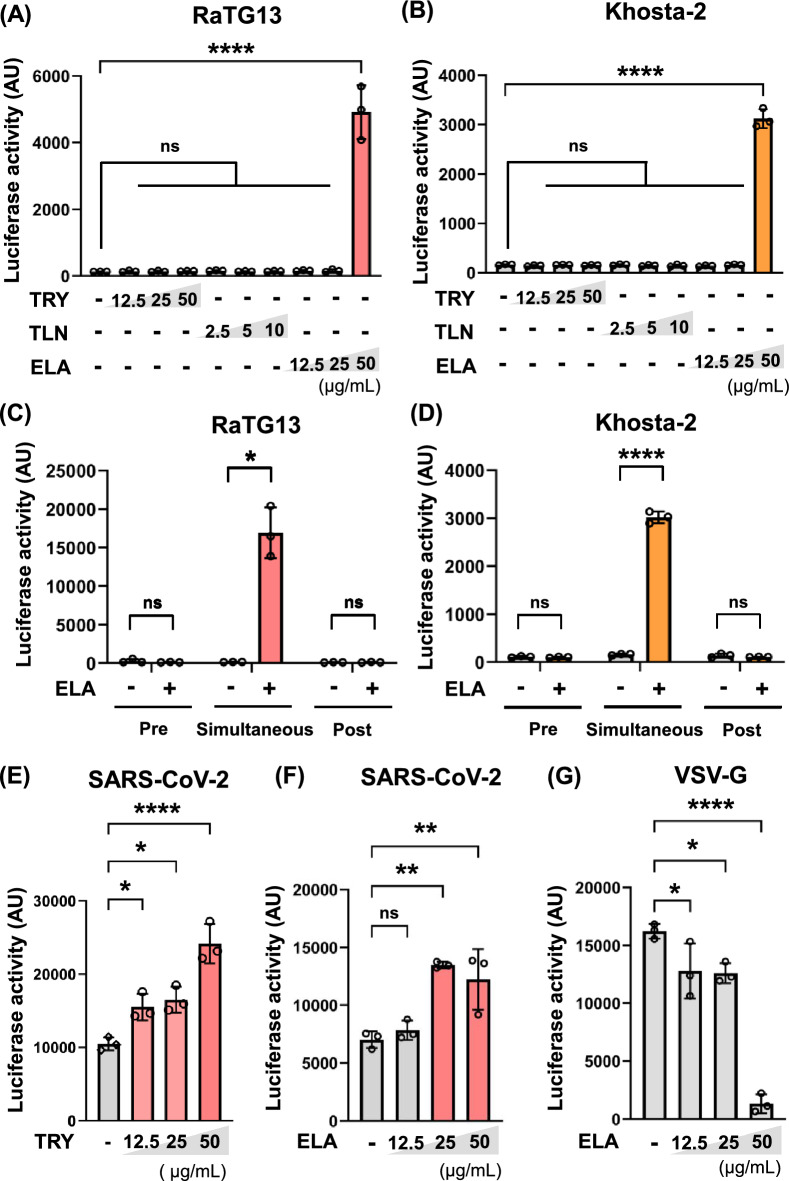
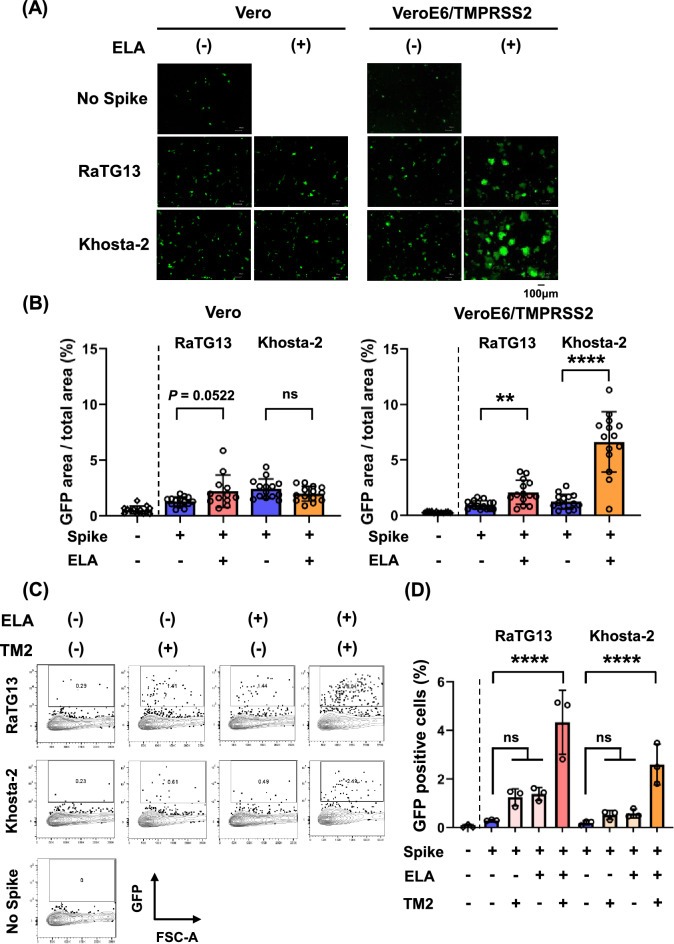
Although numerous sarbecoviruses have been identified in bats, but most lack the ability to infect human cells. Some barriers limit coronavirus zoonosis, including susceptibility to host proteases. Here, we investigated whether exogenous protease treatment can circumvent host restrictions in two severe acute respiratory syndrome (SARS)-related bat coronaviruses. We found that the spike proteins of RaTG13 and Khosta-2, which are sarbecoviruses obtained from horseshoe bats in China and Russia, respectively, facilitated the ACE2-mediated entry of pseudotyped viruses into VeroE6/TMPRSS2 cells following elastase treatment. In contrast, trypsin and thermolysin exhibited no effects. Elastase-enhanced infectivity correlated with increased fusogenicity driven by the cleavage of spike proteins. This process was TMPRSS2-dependent and was inhibited by nafamostat, a TMPRSS2 inhibitor. Additionally, mutation of residue 809 within the S2 subunit of the RaTG13 spike protein (S809D) impaired elastase-induced cleavage and infectivity. Hence, proteolytic processing of the spike protein serves as a restriction to RaTG13 and Khosta-2 infections, which can be overcome by elastase. This suggests that elastase secreted in inflamed tissues during viral infection may increase the zoonotic potential of sarbecoviruses by facilitating human cell entry.
Keywords: Bat SARS-like coronavirus; Elastase; Khosta-2; RaTG13; Spike protein.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures
Similar articles
-
Determinants of susceptibility to SARS-CoV-2 infection in murine ACE2.J Virol. 2025 Jun 17;99(6):e0054325. doi: 10.1128/jvi.00543-25. Epub 2025 May 12. J Virol. 2025. PMID: 40353671 Free PMC article.
-
The ACE2 Receptor from Common Vampire Bat (Desmodus rotundus) and Pallid Bat (Antrozous pallidus) Support Attachment and Limited Infection of SARS-CoV-2 Viruses in Cell Culture.Viruses. 2025 Mar 31;17(4):507. doi: 10.3390/v17040507. Viruses. 2025. PMID: 40284950 Free PMC article.
-
The furin cleavage site is required for pathogenesis, but not transmission, of SARS-CoV-2.J Virol. 2025 Jul 22;99(7):e0046725. doi: 10.1128/jvi.00467-25. Epub 2025 Jun 10. J Virol. 2025. PMID: 40492735 Free PMC article.
-
From Bat to Worse: The Pivotal Role of Bats for Viral Zoonosis.Microb Biotechnol. 2025 Jul;18(7):e70190. doi: 10.1111/1751-7915.70190. Microb Biotechnol. 2025. PMID: 40619741 Free PMC article. Review.
-
Receptor Binding for the Entry Mechanisms of SARS-CoV-2: Insights from the Original Strain and Emerging Variants.Viruses. 2025 May 10;17(5):691. doi: 10.3390/v17050691. Viruses. 2025. PMID: 40431702 Free PMC article. Review.
References
-
- Li, W. et al. Bats are natural reservoirs of SARS-like coronaviruses. Science310, 676–679 (2005). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
