

Systematic screen uncovers regulator contributions to chemical cues in Escherichia coli
- PMID: 40694573
- PMCID: PMC12282887
- DOI: 10.1371/journal.pbio.3003260
Systematic screen uncovers regulator contributions to chemical cues in Escherichia coli
Abstract
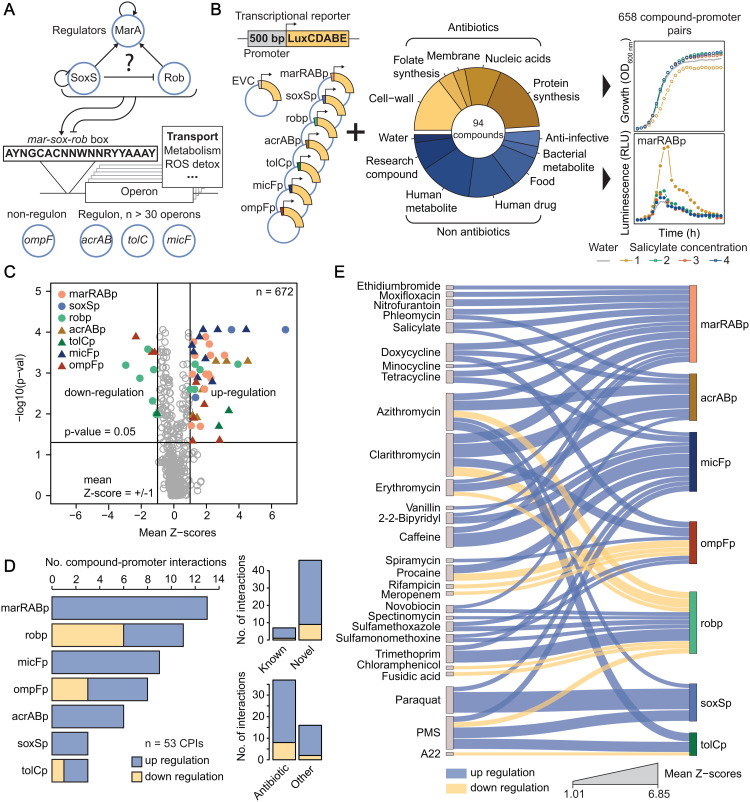
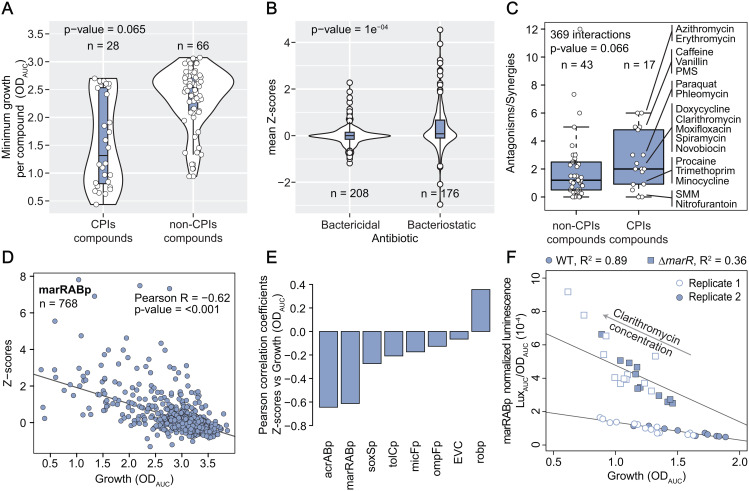
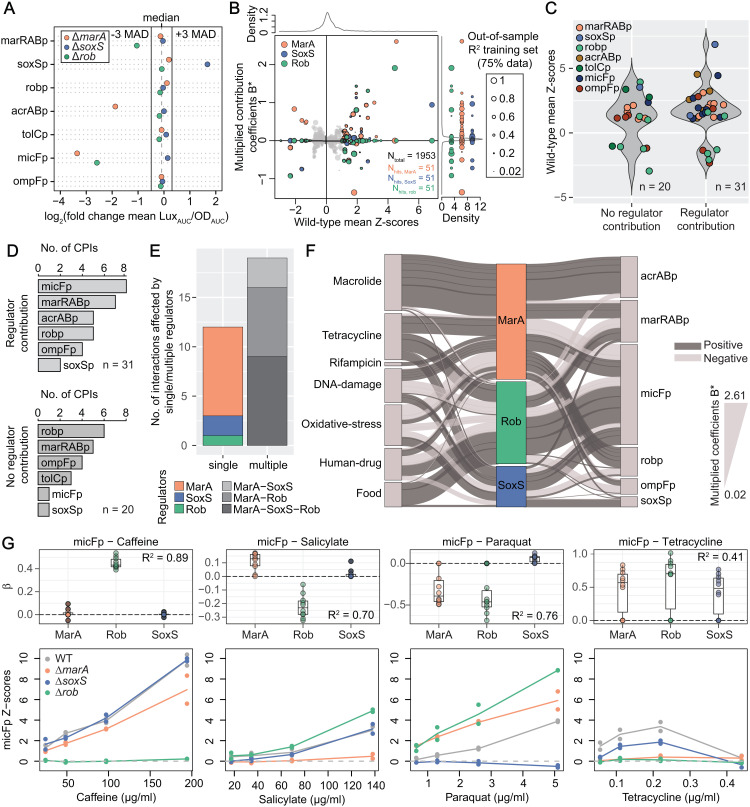
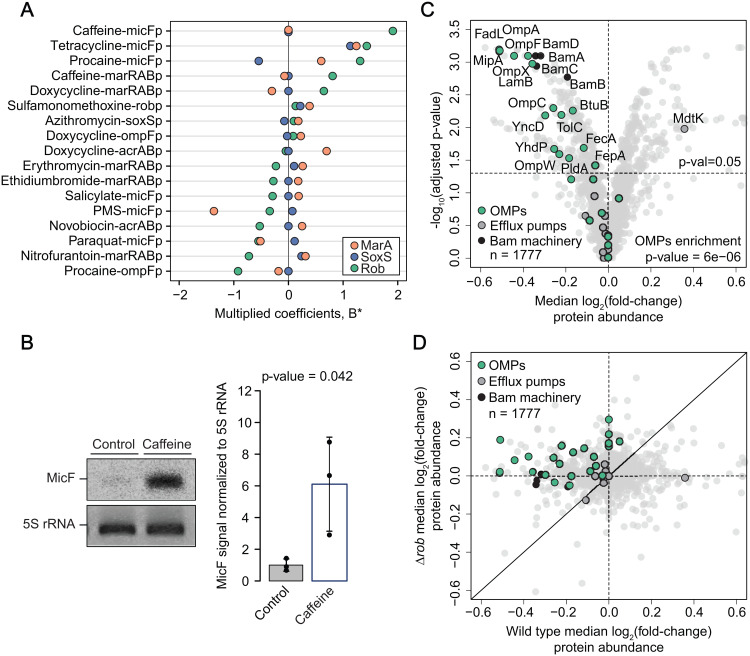
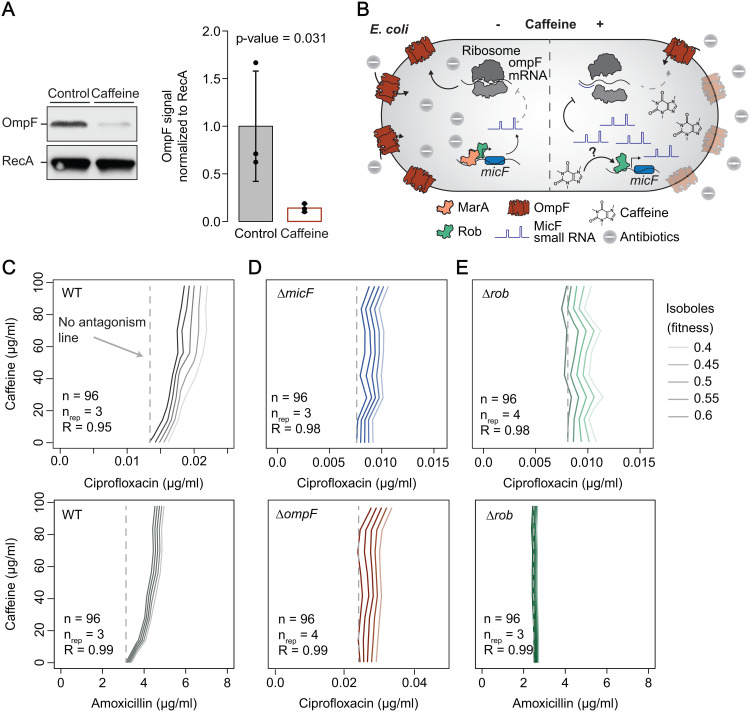
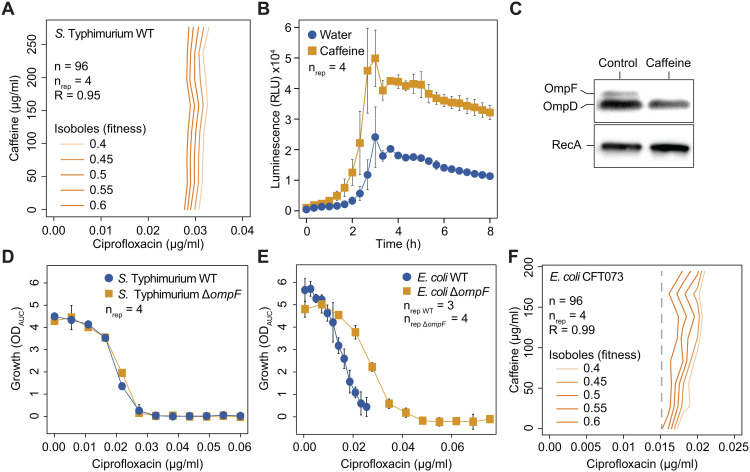
In Gram-negative bacteria, the uptake and export of a wide range of molecules, including antibiotics, is facilitated by porins and efflux pumps. Because of their role in regulating small molecule permeability of the outer and inner membrane, these transport machineries are tightly regulated at the transcriptional and post-transcriptional levels. However, regulation of transport by external chemical cues remains poorly understood. Here we investigated transcriptional regulation of three prominent transporter genes in Escherichia coli across 94 defined chemical cues, and simultaneously mapped the contributions of the key regulators MarA, SoxS and Rob to promoter activity. One third of all tested compounds triggered transcriptional changes, the majority of which were previously unknown. Importantly, we exposed main drivers of transport control in E. coli, e.g., bacteriostatic but not bactericidal antibiotics trigger the expression of efflux pumps, and Rob contributes to ~1/3 of all measured transcriptional changes, thereby emerging as a more prominent regulator of transport than previously thought. We showcase the potential of our resource by elucidating the molecular mechanism of antibiotic antagonisms with widely consumed caffeine in E. coli. Altogether, our analysis provides a quantitative overview of how different regulators orchestrate the transcriptional response of major transport determinants to environmental chemical cues.
Copyright: © 2025 Binsfeld et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
