

Visomitin Attenuates Pathological Bone Loss by Reprogramming Osteoclast Metabolism via the STAT3/LDHB Axis
- PMID: 40698330
- PMCID: PMC12280330
- DOI: 10.34133/research.0784
Visomitin Attenuates Pathological Bone Loss by Reprogramming Osteoclast Metabolism via the STAT3/LDHB Axis
Abstract
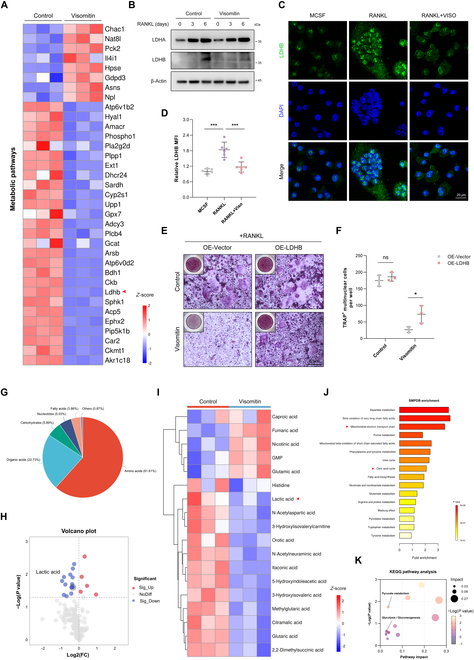
A persistently substantial energy demand and metabolic reprogramming endure throughout the entire course of osteoclastogenesis, accompanied by an intensified oxidative stress. Hence, balancing cellular energy metabolism and maintaining redox homeostasis offer potential for coordinating osteoclastogenesis and bone loss in pathological conditions. In the present study, we have discovered Visomitin, a novel antioxidant that specifically targets mitochondria, which efficiently decreases intracellular reactive oxygen species (ROS) levels, inhibits osteoclastogenesis, and impairs the function of bone resorption. Mechanistically, Visomitin directly targets signal transducer and activator of transcription 3 (STAT3), leading to the inhibition of its transcriptional activity and modulation of lactate dehydrogenase B (LDHB) expression levels, consequently triggering metabolic reprogramming and exerting antagonistic effects on osteoclasts. Furthermore, administration of Visomitin demonstrates marked protective effects against pathological bone loss in vivo. Given its established clinical safety profile in ophthalmologic applications, Visomitin emerges as a promising anti-resorptive agent for clinical translation. This study also unveils the STAT3/LDHB axis as a critical nexus linking mitochondrial redox regulation to osteoclast metabolism, providing a novel therapeutic strategy for osteoclast-driven bone diseases.
Copyright © 2025 Putao Yuan et al.
Conflict of interest statement
Competing interests: The authors declare that they have no competing interests.
Figures





Similar articles
-
Role of Pink1 in Regulating Osteoclast Differentiation during Periodontitis.J Dent Res. 2025 Jul;104(7):753-762. doi: 10.1177/00220345251315723. Epub 2025 Mar 12. J Dent Res. 2025. PMID: 40075549
-
Downregulation of the metalloproteinases ADAM10 or ADAM17 promotes osteoclast differentiation.Cell Commun Signal. 2024 Jun 11;22(1):322. doi: 10.1186/s12964-024-01690-y. Cell Commun Signal. 2024. PMID: 38863060 Free PMC article.
-
Downregulation of LATS1/2 Drives Endothelial Senescence-Associated Stemness (SAS) and Atherothrombotic Lesion Formation.bioRxiv [Preprint]. 2025 Jun 21:2025.06.19.660635. doi: 10.1101/2025.06.19.660635. bioRxiv. 2025. PMID: 40667385 Free PMC article. Preprint.
-
Nrf2 signaling pathway: focus on oxidative stress in osteoporosis.Osteoporos Int. 2025 Jul 15. doi: 10.1007/s00198-025-07592-0. Online ahead of print. Osteoporos Int. 2025. PMID: 40663116 Review.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
References
-
- Veis DJ, O’Brien CA. Osteoclasts, master sculptors of bone. Annu Rev Pathol. 2023;18:257–281. - PubMed
-
- Singer FR, Leach RJ. Bone: Do all Paget disease risk genes incriminate the osteoclast? Nat Rev Rheumatol. 2010;6(9):502–503. - PubMed
-
- Walsh NC, Gravallese EM. Bone remodeling in rheumatic disease: A question of balance. Immunol Rev. 2010;233(1):301–312. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
