

Hepatitis B surface antigen is upregulated by HIV Tat in an HIV-hepatitis B virus co-infection model system
- PMID: 40698826
- PMCID: PMC12403812
- DOI: 10.1128/spectrum.00809-25
Hepatitis B surface antigen is upregulated by HIV Tat in an HIV-hepatitis B virus co-infection model system
Abstract
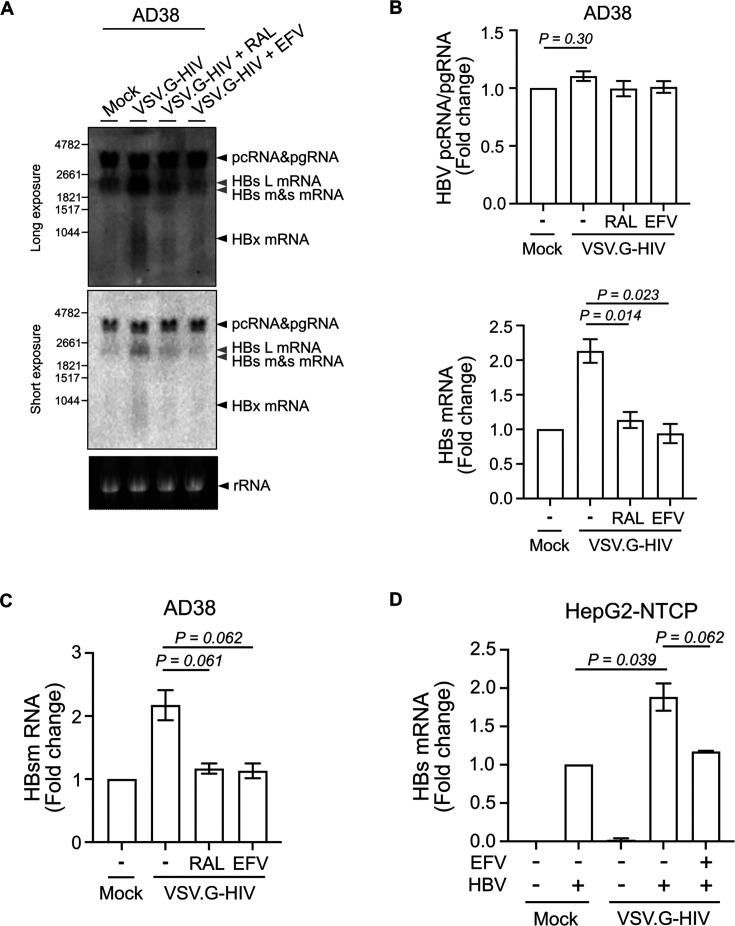
People with human immunodeficiency virus-hepatitis B virus (HIV-HBV) co-infection have faster rates of liver disease progression and an increase in hepatocellular carcinoma compared to people with HBV mono-infection. Given that HIV can infect multiple cells in the liver, including hepatocytes, we hypothesized that HIV will impact HBV replication through HIV viral proteins that can impact HBV replication either directly or indirectly, via effects on cellular pathways. Following infection of sodium taurocholate co-transporting polypeptide (NTCP)-expressing HepG2 cells with HBV and vesicular stomatitis virus G protein (VSV.G)-pseudotyped HIV, we found that productive HIV infection led to a twofold upregulation of HBV surface (HBs) mRNA and a marked increase in intracellular production and cellular retention of HBs antigen (HBsAg). Overexpression of HIV Tat protein, but not other HIV proteins, by DNA plasmid transfection in the HBV-producing cell line AD38 significantly stimulated HBs mRNA expression. This could be rescued by CDK9 inhibition with BAY-1251152. This study provides new insights into the mechanisms by which HIV directly impacts HBV replication and has implications for understanding adverse liver outcomes in people living with HIV and HBV.IMPORTANCEPeople with both human immunodeficiency virus (HIV) and hepatitis B virus (HBV) face faster liver disease progression and a higher risk of liver cancer than those with HBV alone. This study investigated how HIV affects HBV replication in liver cells and found that HIV infection increases the production of a key HBV surface protein (HBsAg) by enhancing the expression of its gene (HBs). This effect is driven by the HIV Tat protein. Notably, blocking the CDK9 pathway prevented this increase, suggesting a possible explanation for the adverse liver outcomes in co-infected individuals. Our findings have implications for interventions aiming to cure HIV through latency reversal, as these interventions can specifically increase the Tat protein. Future exploratory treatment strategies, such as Tat inhibitors, could play a role in the management of people with HIV and HBV at high risk of liver disease.
Keywords: CDK9; HBsAg; Tat; co-infection; hepatitis B virus; hepatocyte; human immunodeficiency virus; liver.
Conflict of interest statement
S.R.L. has received funding from the National Health and Medical Research Council of Australia (NHMRC), the National Institutes of Health (NIH), amfAR, The Australian Centre for HIV and Hepatitis Research, Melbourne HIV Cure Consortium, Gilead Sciences, Merck, ViiV, Abbvie, and Gilead outside the submitted work. F.J.R. receives institutional support as a coinvestigator and is subcontracted by the Peter MacCallum Cancer Centre for an investigator-initiated trial, which receives funding support from Sanofi/Regeneron Pharmaceuticals.
Figures



Similar articles
-
The loss of hepatitis B virus receptor NTCP/SLC10A1 in human liver cancer cells is due to epigenetic silencing.J Virol. 2024 Oct 22;98(10):e0118724. doi: 10.1128/jvi.01187-24. Epub 2024 Sep 19. J Virol. 2024. PMID: 39297647 Free PMC article.
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Hepatitis B immunoglobulin during pregnancy for prevention of mother-to-child transmission of hepatitis B virus.Cochrane Database Syst Rev. 2017 Feb 11;2(2):CD008545. doi: 10.1002/14651858.CD008545.pub2. Cochrane Database Syst Rev. 2017. PMID: 28188612 Free PMC article.
-
Prevalence, Risk Factors, and Clinical Profiles of Hepatitis D Virus in Nigeria: A Systematic Review, 2009-2024.Viruses. 2024 Oct 31;16(11):1723. doi: 10.3390/v16111723. Viruses. 2024. PMID: 39599836 Free PMC article.
-
Adefovir dipivoxil and pegylated interferon alfa-2a for the treatment of chronic hepatitis B: a systematic review and economic evaluation.Health Technol Assess. 2006 Aug;10(28):iii-iv, xi-xiv, 1-183. doi: 10.3310/hta10280. Health Technol Assess. 2006. PMID: 16904047
References
-
- Platt L, French CE, McGowan CR, Sabin K, Gower E, Trickey A, McDonald B, Ong J, Stone J, Easterbrook P, Vickerman P. 2020. Prevalence and burden of HBV co-infection among people living with HIV: a global systematic review and meta-analysis. J Viral Hepat 27:294–315. doi: 10.1111/jvh.13217 - DOI - PMC - PubMed
-
- Boyd A, Gozlan J, Maylin S, Delaugerre C, Peytavin G, Girard PM, Zoulim F, Lacombe K. 2014. Persistent viremia in human immunodeficiency virus/hepatitis B coinfected patients undergoing long-term tenofovir: virological and clinical implications. Hepatology 60:497–507. doi: 10.1002/hep.27182 - DOI - PubMed
-
- Klein MB, Althoff KN, Jing Y, Lau B, Kitahata M, Lo Re V 3rd, Kirk GD, Hull M, Kim HN, Sebastiani G, et al. 2016. Risk of end-stage liver disease in HIV-viral hepatitis coinfected persons in North America from the early to modern antiretroviral therapy eras. Clin Infect Dis 63:1160–1167. doi: 10.1093/cid/ciw531 - DOI - PMC - PubMed
-
- Dezanet LNC, Kassime R, Miailhes P, Lascoux-Combe C, Chas J, Maylin S, Gabassi A, Rougier H, Delaugerre C, Lacombe K, Boyd A. 2021. Effect of viral replication and liver fibrosis on all-cause mortality in HIV/HBV coinfected individuals: a retrospective analysis of a 15-year longitudinal cohort. MedRxiv. doi: 10.1101/2021.04.13.21255432 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
