

The N-Linked Glycosylation Site N201 in eel Lutropin/Choriogonadotropin Receptor Is Uniquely Indispensable for cAMP Responsiveness and Receptor Surface Loss, but Not pERK1/2 Activity
- PMID: 40699744
- PMCID: PMC12110046
- DOI: 10.3390/cimb47050345
The N-Linked Glycosylation Site N201 in eel Lutropin/Choriogonadotropin Receptor Is Uniquely Indispensable for cAMP Responsiveness and Receptor Surface Loss, but Not pERK1/2 Activity
Abstract
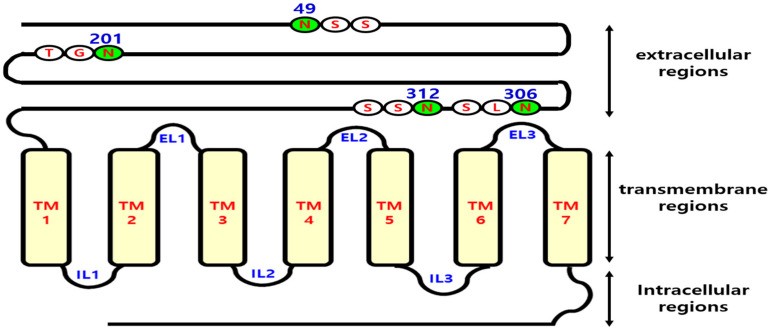
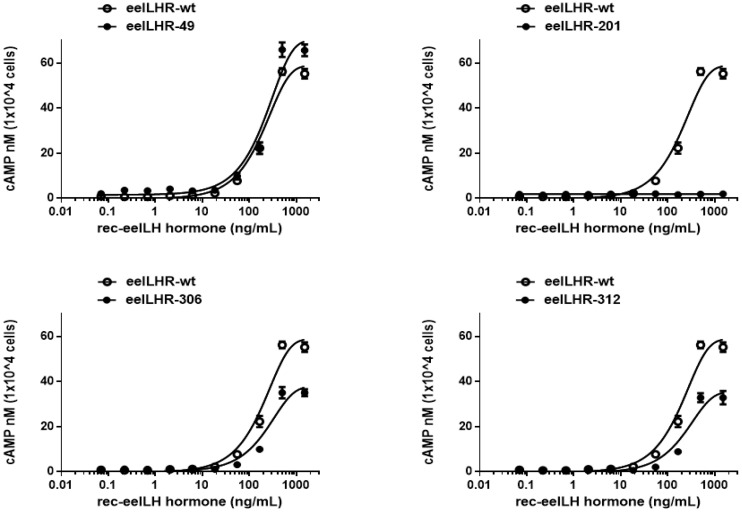
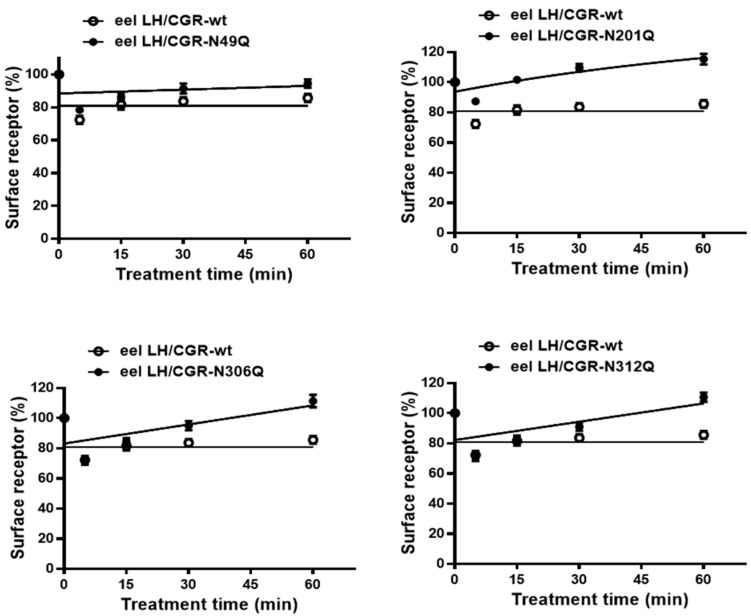
The seven transmembrane-spanning lutropin/chorionic gonadotropin receptors (LH/CGRs) trigger extracellular signal-related kinases (ERK1/2) via a noticeable network dependent on either G protein (Gαs) or β-arrestins. LH/CGRs are highly conserved, with the largest region within the transmembrane helices and common N-glycosylation sites in the extracellular domain. We aimed to determine the glycosylation sites that play crucial roles in cAMP and pERK1/2 regulation by constructing four mutants (N49Q, N201Q, N306Q, and N312Q). The cAMP response in cells expressing the N201Q mutant was completely impaired, despite high-dose agonist treatment. The cell-surface expression level was lowest in transiently transfected cells, but normal surface loss of the receptor occurred in cells expressing the wild-type and other mutant proteins. However, the N201Q mutant was only slightly reduced after 5 min of agonist stimulation. All mutants showed a peak in cAMP signaling 5 min after stimulation with a pERK1/2 agonist. Of note, cAMP activity was completely impaired in the N201Q mutant; however, this mutant still displayed a pERK1/2 response. These data show that the specific N-linked glycosylation site in eel LH/CGR is clearly distinguished by its differential responsiveness to cAMP signaling and pERK1/2 activity. Thus, we suggest that the cAMP and pERK1/2 signaling pathways involving eel LH/CGRs represent pleiotropic signal transduction induced by agonist treatment.
Keywords: N-linked glycosylation; cAMP responsive; eel LH/CGR; pERK1/2 activity.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures






Similar articles
-
Functional Divergence for N-Linked Glycosylation Sites in Equine Lutropin/Choriogonadotropin Receptors.Curr Issues Mol Biol. 2025 Jul 25;47(8):590. doi: 10.3390/cimb47080590. Curr Issues Mol Biol. 2025. PMID: 40864744 Free PMC article.
-
Signal Transduction of Constitutive Activating and Inactivating Eel Lutropin/Choriogonadotropin Receptor (Eel LH/CGR) Mutants by Recombinant Equine Chorionic Gonadotropin (Rec-eCG).Dev Reprod. 2024 Dec;28(4):141-151. doi: 10.12717/DR.2024.28.4.141. Epub 2024 Dec 31. Dev Reprod. 2024. PMID: 39845510 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Drugs for preventing postoperative nausea and vomiting in adults after general anaesthesia: a network meta-analysis.Cochrane Database Syst Rev. 2020 Oct 19;10(10):CD012859. doi: 10.1002/14651858.CD012859.pub2. Cochrane Database Syst Rev. 2020. PMID: 33075160 Free PMC article.
-
Pharmacological treatments in panic disorder in adults: a network meta-analysis.Cochrane Database Syst Rev. 2023 Nov 28;11(11):CD012729. doi: 10.1002/14651858.CD012729.pub3. Cochrane Database Syst Rev. 2023. PMID: 38014714 Free PMC article.
References
-
- Kara E., Crepieux P., Gauthier C., Martinat N., Piketty V., Guillou F., Reiter E. A phosphorylation cluster of five serine and threonine residues in the C-terminus of the follicle-stimulating hormone receptor is important for desensitization but not for β-arrestin-mediated ERK activation. Mol. Endocrinol. 2006;20:3014–3026. doi: 10.1210/me.2006-0098. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
