

Stochastic cell-intrinsic stem cell decisions control colony growth in planarians
- PMID: 40699888
- PMCID: PMC12286600
- DOI: 10.7554/eLife.100885
Stochastic cell-intrinsic stem cell decisions control colony growth in planarians
Abstract
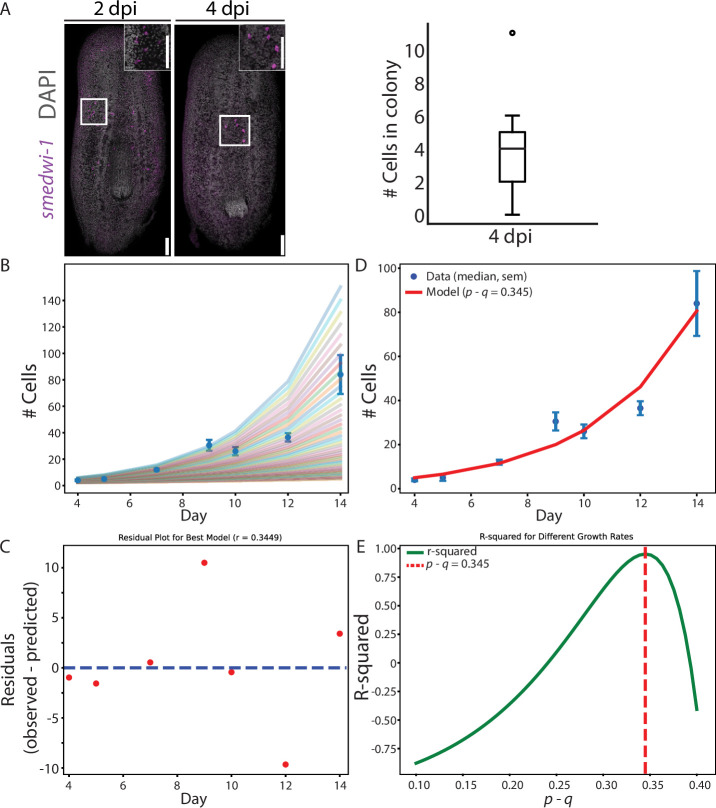
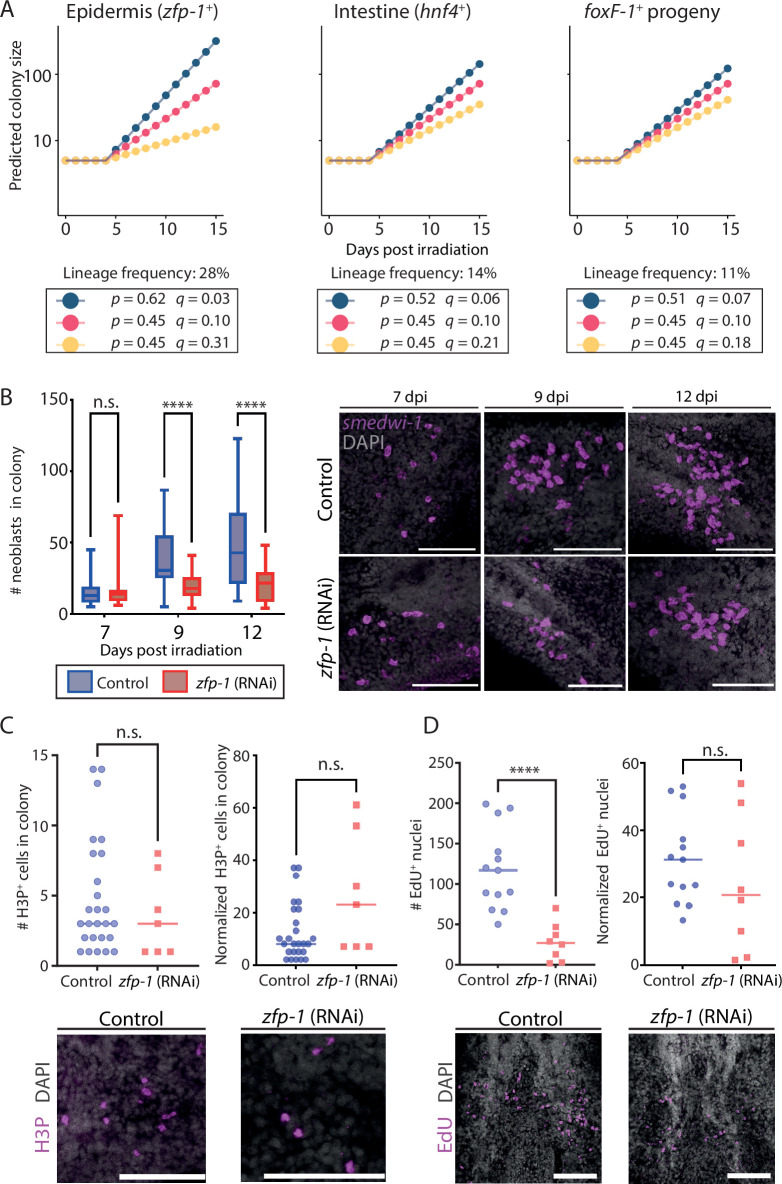
Stem cells contribute to organismal homeostasis by balancing division, self-renewal, and differentiation. Elucidating the strategies by which stem cells achieve this balance is critical for understanding homeostasis and for addressing pathogenesis associated with the disruption of this balance (e.g. cancer). Planarians, highly regenerative flatworms, use pluripotent stem cells called neoblasts to maintain and regrow organs. A single neoblast can rescue an entire animal depleted from stem cells and regenerate all cell lineages. How neoblast differentiation and clonal expansion are governed to produce all the required cell types remains unclear. Here, we integrated experimental and computational approaches to develop a quantitative model revealing basic principles of clonal growth of individual neoblasts. By experimentally suppressing differentiation to major lineages, we elucidated the interplay between colony growth and lineage decisions. Our findings suggest that neoblasts select their progenitor lineage based on a cell-intrinsic fate distribution. Arresting differentiation into specific lineages disrupts neoblast proliferative capacity without inducing compensatory expression of other lineages. Our analysis of neoblast colonies is consistent with a cell-intrinsic decision model that can operate without memory or communication between neoblasts. This simple cell fate decision process breaks down in homeostasis, likely because of the activity of feedback mechanisms. Our findings uncover essential principles of stem cell regulation in planarians, which are distinct from those observed in many vertebrate models. These mechanisms enable robust production of diverse cell types and facilitate regeneration of missing tissues.
Keywords: Schmidtea mediterranea; cell fate decision; developmental biology; neoblast; planarian; regeneration; regenerative medicine; stem cell; stem cells; systems modeling.
© 2024, Frankovits et al.
Conflict of interest statement
TF, PC, YY, SD, OW No competing interests declared
Figures







Update of
- doi: 10.1101/2024.02.26.582030
- doi: 10.7554/eLife.100885.1
- doi: 10.7554/eLife.100885.2
Similar articles
-
Derivation of trophoblast stem cells from naïve human pluripotent stem cells.Elife. 2020 Feb 12;9:e52504. doi: 10.7554/eLife.52504. Elife. 2020. PMID: 32048992 Free PMC article.
-
The DNA methyltransferase DMAP1 is required for tissue maintenance and planarian regeneration.Dev Biol. 2024 Dec;516:196-206. doi: 10.1016/j.ydbio.2024.08.007. Epub 2024 Aug 22. Dev Biol. 2024. PMID: 39179016 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Fabricating mice and dementia: opening up relations in multi-species research.In: Jenkins N, Jack-Waugh A, Ritchie L, editors. Multi-Species Dementia Studies. Bristol (UK): Bristol University Press; 2025 Feb 25. Chapter 2. In: Jenkins N, Jack-Waugh A, Ritchie L, editors. Multi-Species Dementia Studies. Bristol (UK): Bristol University Press; 2025 Feb 25. Chapter 2. PMID: 40690569 Free Books & Documents. Review.
-
Stem cell insights into human trophoblast lineage differentiation.Hum Reprod Update. 2016 Dec;23(1):77-103. doi: 10.1093/humupd/dmw026. Epub 2016 Sep 2. Hum Reprod Update. 2016. PMID: 27591247
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
