

Parabrachial Foxp2-expressing neurons are necessary for sustaining core body temperature in the cold
- PMID: 40703448
- PMCID: PMC12283563
- DOI: 10.1016/j.isci.2025.112764
Parabrachial Foxp2-expressing neurons are necessary for sustaining core body temperature in the cold
Abstract
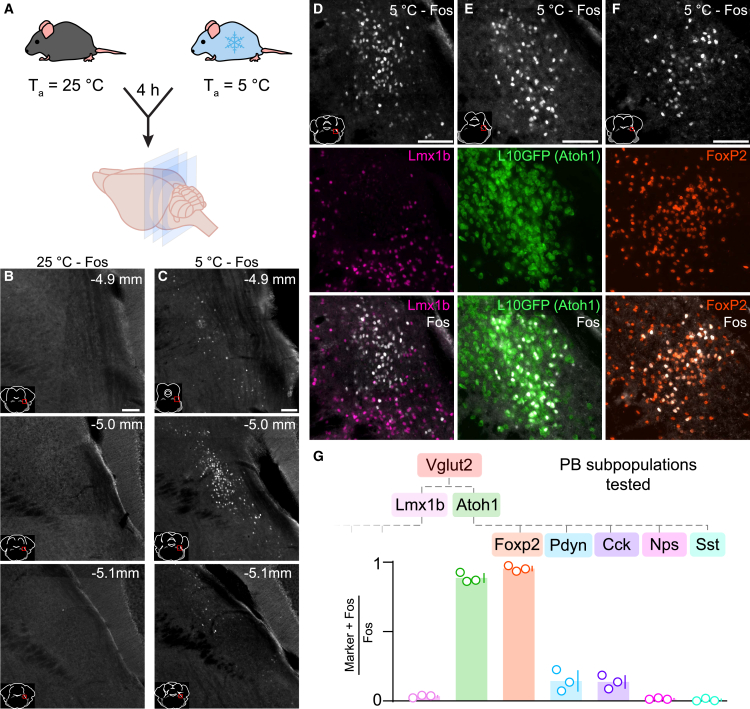
Cold environmental temperature is a threat to survival. Sustaining core body temperature in the cold requires a dynamic set of adaptive responses known as "cold defense," but the neural circuitry orchestrating these responses remains unclear. We identified a cluster of Atoh1-derived, Foxp2-expressing glutamatergic neurons in the lateral parabrachial nucleus (PB) that are activated by exposing mice to cold environmental temperature. Eliminating Foxp2-expressing PB neurons caused body temperature to plummet in the cold. Mice lacking these neurons had normal wakefulness, movement and appetite at room temperature, and their autonomic cold-defense responses remained intact. However, these mice had reduced metabolism and locomotor activity in the cold, and thermal discrimination was impaired. Our results indicate that thermosensory information relayed through Foxp2-expressing PB neurons is essential for sensing and surviving a cold environment.
Keywords: Biological sciences; Natural sciences; Neuroscience; Physiology; Sensory neuroscience; Systems neuroscience.
© 2025 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures







Similar articles
-
Efferent projections of Vglut2, Foxp2, and Pdyn parabrachial neurons in mice.J Comp Neurol. 2021 Mar;529(4):657-693. doi: 10.1002/cne.24975. Epub 2020 Sep 21. J Comp Neurol. 2021. PMID: 32621762 Free PMC article.
-
Molecular ontology of the parabrachial nucleus.J Comp Neurol. 2022 Jul;530(10):1658-1699. doi: 10.1002/cne.25307. Epub 2022 Feb 21. J Comp Neurol. 2022. PMID: 35134251 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Active body surface warming systems for preventing complications caused by inadvertent perioperative hypothermia in adults.Cochrane Database Syst Rev. 2016 Apr 21;4(4):CD009016. doi: 10.1002/14651858.CD009016.pub2. Cochrane Database Syst Rev. 2016. PMID: 27098439 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
References
-
- Dhaka A., Murray A.N., Mathur J., Earley T.J., Petrus M.J., Patapoutian A. TRPM8 is required for cold sensation in mice. Neuron. 2007;54:371–378. - PubMed
-
- Bautista D.M., Siemens J., Glazer J.M., Tsuruda P.R., Basbaum A.I., Stucky C.L., Jordt S.-E., Julius D. The menthol receptor TRPM8 is the principal detector of environmental cold. Nature. 2007;448:204–208. - PubMed
-
- Craig A.D., Krout K., Andrew D. Quantitative response characteristics of thermoreceptive and nociceptive lamina I spinothalamic neurons in the cat. J. Neurophysiol. 2001;86:1459–1480. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
