

Aldolase A accelerates hepatocarcinogenesis by refactoring c-Jun transcription
- PMID: 40708574
- PMCID: PMC12284681
- DOI: 10.1016/j.jpha.2024.101169
Aldolase A accelerates hepatocarcinogenesis by refactoring c-Jun transcription
Abstract
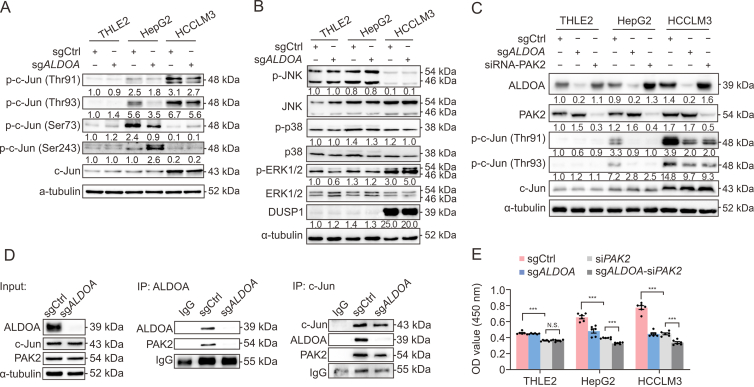
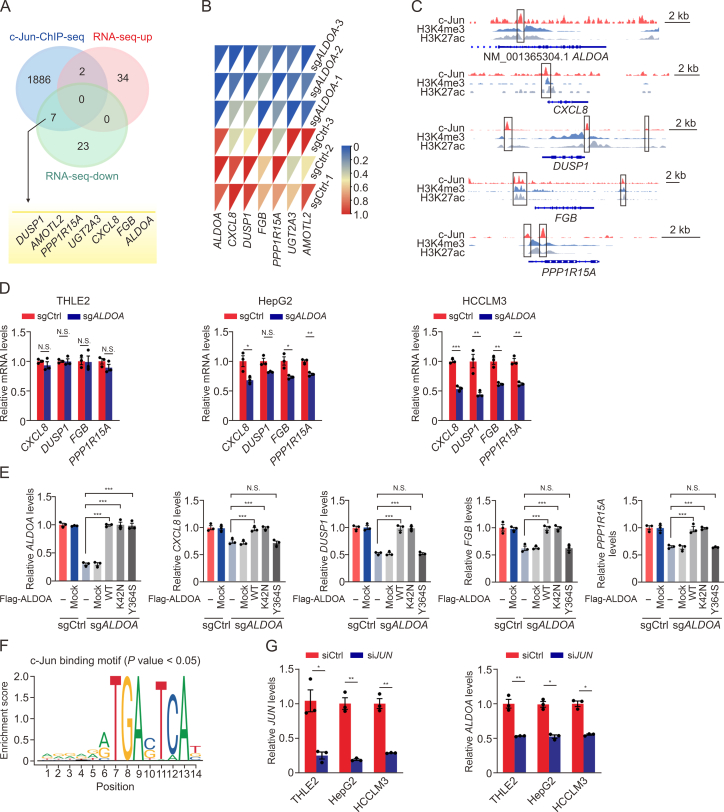
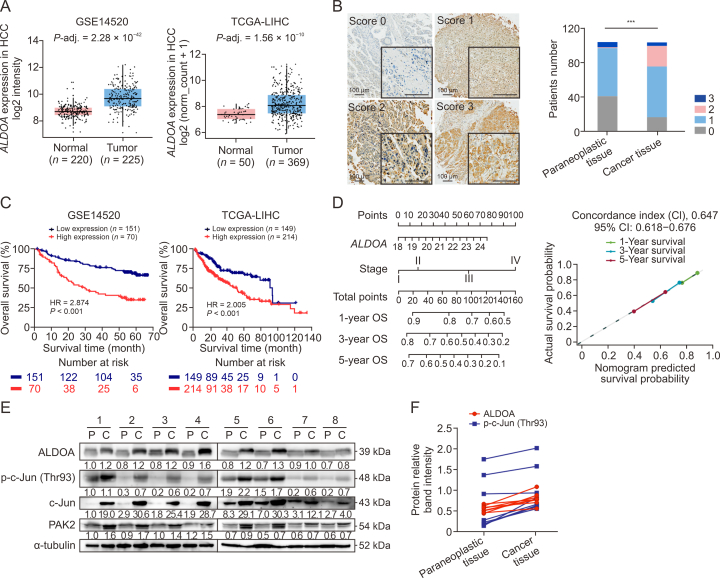
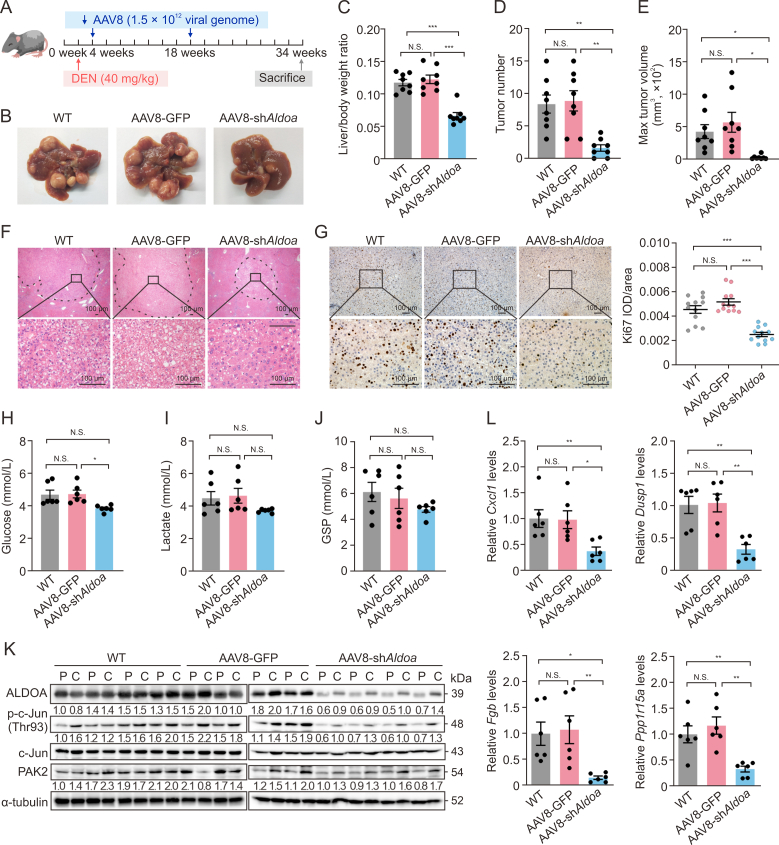
Hepatocellular carcinoma (HCC) expresses abundant glycolytic enzymes and displays comprehensive glucose metabolism reprogramming. Aldolase A (ALDOA) plays a prominent role in glycolysis; however, little is known about its role in HCC development. In the present study, we aim to explore how ALDOA is involved in HCC proliferation. HCC proliferation was markedly suppressed both in vitro and in vivo following ALDOA knockout, which is consistent with ALDOA overexpression encouraging HCC proliferation. Mechanistically, ALDOA knockout partially limits the glycolytic flux in HCC cells. Meanwhile, ALDOA translocated to nuclei and directly interacted with c-Jun to facilitate its Thr93 phosphorylation by P21-activated protein kinase; ALDOA knockout markedly diminished c-Jun Thr93 phosphorylation and then dampened c-Jun transcription function. A crucial site Y364 mutation in ALDOA disrupted its interaction with c-Jun, and Y364S ALDOA expression failed to rescue cell proliferation in ALDOA deletion cells. In HCC patients, the expression level of ALDOA was correlated with the phosphorylation level of c-Jun (Thr93) and poor prognosis. Remarkably, hepatic ALDOA was significantly upregulated in the promotion and progression stages of diethylnitrosamine-induced HCC models, and the knockdown of A ldoa strikingly decreased HCC development in vivo. Our study demonstrated that ALDOA is a vital driver for HCC development by activating c-Jun-mediated oncogene transcription, opening additional avenues for anti-cancer therapies.
Keywords: Glycolysis; Hepatocellular carcinoma; Nuclear localization; Transcriptional regulation; c-Jun.
© 2024 The Author(s).
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures





Similar articles
-
mTOR regulates aerobic glycolysis through NEAT1 and nuclear paraspeckle-mediated mechanism in hepatocellular carcinoma.Theranostics. 2022 Apr 24;12(7):3518-3533. doi: 10.7150/thno.72581. eCollection 2022. Theranostics. 2022. PMID: 35547764 Free PMC article.
-
High glucose facilitates hepatocellular carcinoma cell proliferation and invasion via WTAP-mediated HK2 mRNA stability.Mol Cell Biochem. 2025 Jul;480(7):4149-4168. doi: 10.1007/s11010-025-05235-w. Epub 2025 Mar 3. Mol Cell Biochem. 2025. PMID: 40032749
-
Nuclear factor IA-mediated transcriptional regulation of crystallin αB inhibits hepatocellular carcinoma progression.Mol Clin Oncol. 2025 Jun 20;23(2):72. doi: 10.3892/mco.2025.2867. eCollection 2025 Aug. Mol Clin Oncol. 2025. PMID: 40599718 Free PMC article.
-
A systematic review of p53 regulation of oxidative stress in skeletal muscle.Redox Rep. 2018 Dec;23(1):100-117. doi: 10.1080/13510002.2017.1416773. Epub 2018 Jan 3. Redox Rep. 2018. PMID: 29298131 Free PMC article.
-
C-Jun Transcription Factor Oncogenic Activation in Oral Carcinoma.Maedica (Bucur). 2024 Jun;19(2):350-354. doi: 10.26574/maedica.2024.19.2.350. Maedica (Bucur). 2024. PMID: 39188842 Free PMC article. Review.
References
-
- Warburg O. On the origin of cancer cells. Science. 1956;123:309–314. - PubMed
-
- Garber K. Energy deregulation: Licensing tumors to grow. Science. 2006;312:1158–1159. - PubMed
-
- Sun J., Ding J., Shen Q., et al. Decreased propionyl-CoA metabolism facilitates metabolic reprogramming and promotes hepatocellular carcinoma. J. Hepatol. 2023;78:627–642. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
