

Fibroblasts activated by miRs-185-5p, miR-652-5p, and miR-1246 shape the tumor microenvironment in triple-negative breast cancer via PATZ1 downregulation
- PMID: 40711489
- PMCID: PMC12297133
- DOI: 10.1007/s00018-025-05781-y
Fibroblasts activated by miRs-185-5p, miR-652-5p, and miR-1246 shape the tumor microenvironment in triple-negative breast cancer via PATZ1 downregulation
Abstract
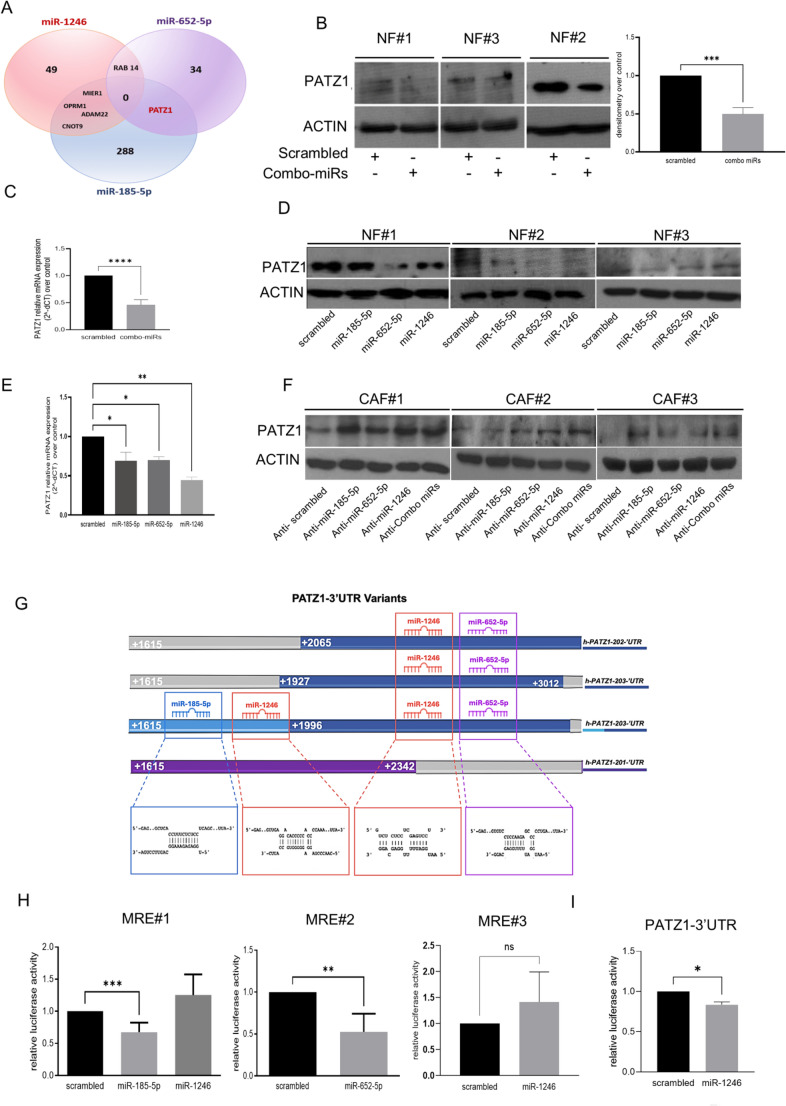
The intricate interplay between epithelial and fibroblast cells within the tumor microenvironment plays a crucial role in driving triple-negative breast cancer progression. This crosstalk involves the exchange of various signaling molecules, including growth factors, cytokines, extracellular matrix components, and extracellular vesicles. Recently, we demonstrated that triple-negative breast cancer extracellular vesicles carry and release a specific combination of miRs, including miR-185-5p, miR-652-5p, and miR-1246 (from here on, referred as combo-miRs), into normal fibroblasts, effectively reprogramming them into cancer-associated fibroblasts. Here, we show that the conditioned medium from the fibroblasts activated by combo-miRs exerts a pro-tumorigenic effect on epithelial cells, enhancing the viability and migratory potential while driving increased invasiveness in patient-derived breast cancer organoids. A proteomic analysis of conditioned medium from combo-miRs activated fibroblasts revealed 76 significantly upregulated secreted proteins compared to control. Bioinformatic analysis identified the transcriptional factor PATZ1 as a potential regulator of the 12 most highly upregulated proteins. Consistently, in-silico predictions and in vitro experiments confirmed that PATZ1 is a direct target of miR-185-5p and miR-652-5p. The downregulation of PATZ1 by these miRNAs led to increased levels of the secreted proteins in the conditioned medium from combo-miRs activated fibroblasts. Furthermore, the conditioned medium from PATZ1-knockout mesenchymal embryonic fibroblasts and normal fibroblasts with silenced PATZ1 similarly enhanced the migratory potential of MCF10A cells, further supporting the critical role of PATZ1 in regulating tumor-promoting mechanisms. These findings provide valuable insights into the dynamics of the TME in TNBC, highlighting combo-miRs and PATZ1 as promising targets for future therapeutic interventions.
Keywords: CAFs; Extracellular vesicles; Fibroblasts; PATZ1; miRNAs.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: Alessandra Affinito and Sara Verde are employees of AKA Biotech SRL. However, this employment did not influence the design, execution, or interpretation of the research presented in this manuscript. The other authors declare no other competing interests.
Figures



References
-
- Nolan E, Lindeman GJ, Visvader JE (2023) Deciphering breast cancer: from biology to the clinic. Cell 186:1708–1728. 10.1016/j.cell.2023.01.040 - PubMed
-
- Farkas AH, Nattinger AB (2023) Breast Cancer screening and prevention. Ann Intern Med 176:ITC161–ITC. 10.7326/AITC202311210 - PubMed
-
- Patel G, Prince A, Harries M (2024) Advanced Triple-Negative breast Cancer. Semin Oncol Nurs 40:151548. 10.1016/j.soncn.2023.151548 - PubMed
-
- Capuozzo M, Celotto V, Santorsola M, Fabozzi A, Landi L, Ferrara F, Borzacchiello A, Granata V, Sabbatino F, Savarese G, Cascella M, Perri F, Ottaiano A (2023) Emerging treatment approaches for triple-negative breast cancer. Med Oncol 41:5. 10.1007/s12032-023-02257-6 - PubMed
MeSH terms
Substances
Grants and funding
- PNRR-MUR NextGenerationEU cod. CN00000041-/Ministero dell'Università e della Ricerca
- NextGeneration EU PRIN 2022 (M5LBKP)/Ministero dell'Università e della Ricerca
- European Union NextGeneration-EU PRIN-PNRR (P2022N28FJ)/Ministero dell'Università e della Ricerca
- NextGeneration EU PRIN 2022 (F3ZKJF)/Ministero dell'Università e della Ricerca
- NextGeneration EU PRIN 2022 (202279JKW5/Ministero dell'Università e della Ricerca
LinkOut - more resources
Full Text Sources
