

Biallelic BRCA2 variants induce premature ovarian insufficiency by impaired meiotic homologous recombination
- PMID: 40715368
- PMCID: PMC12297597
- DOI: 10.1038/s42003-025-08426-9
Biallelic BRCA2 variants induce premature ovarian insufficiency by impaired meiotic homologous recombination
Abstract
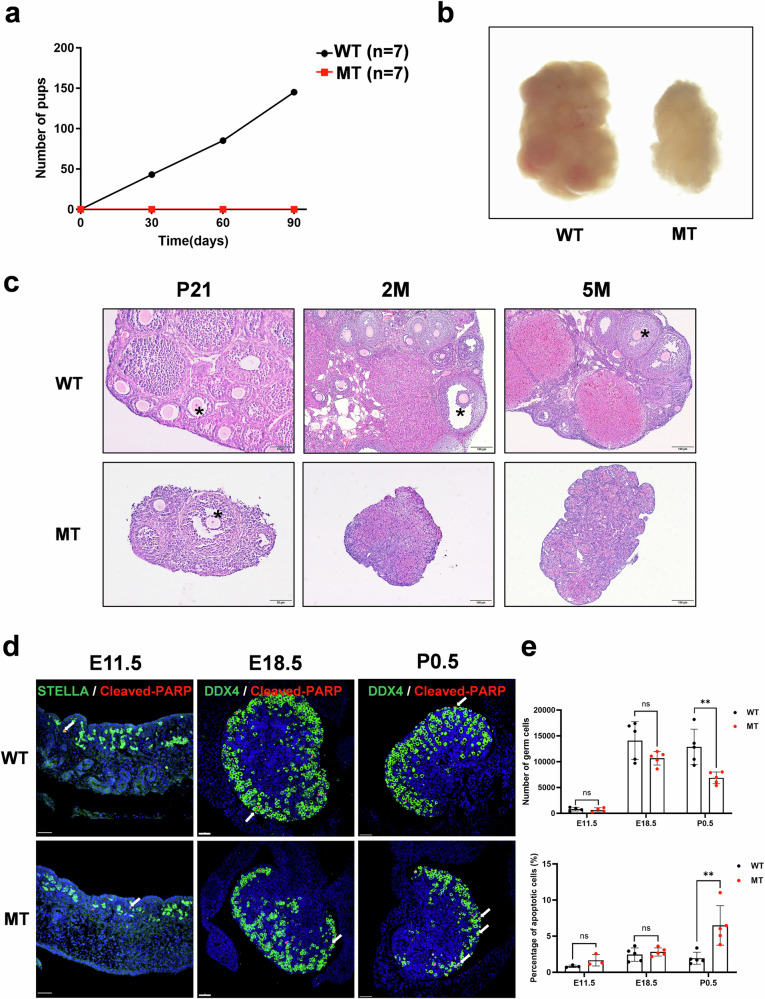
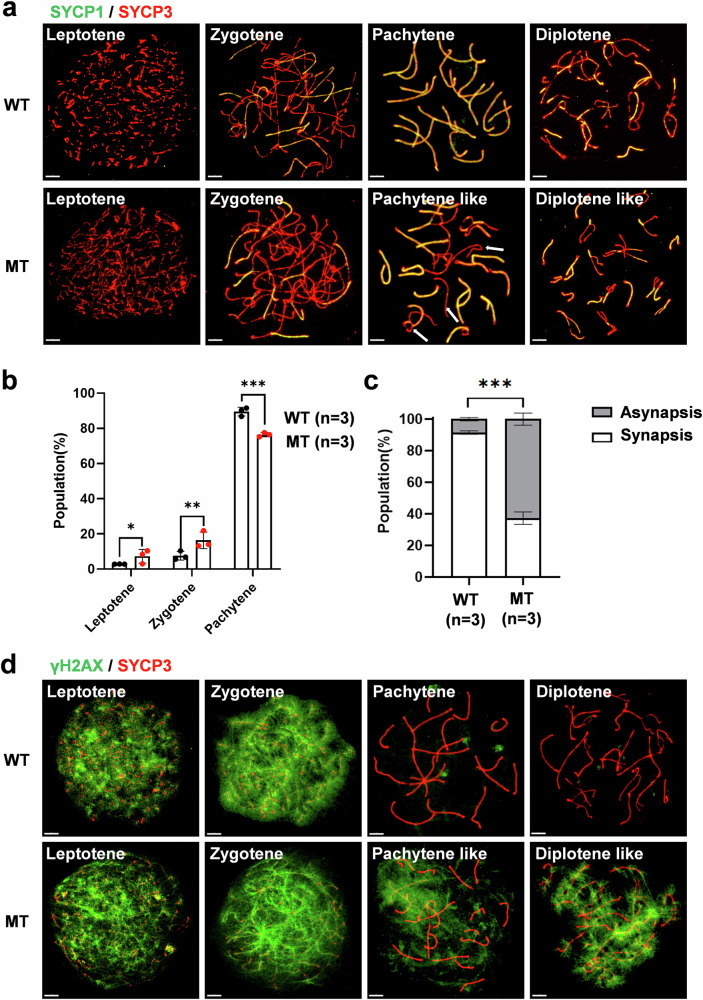
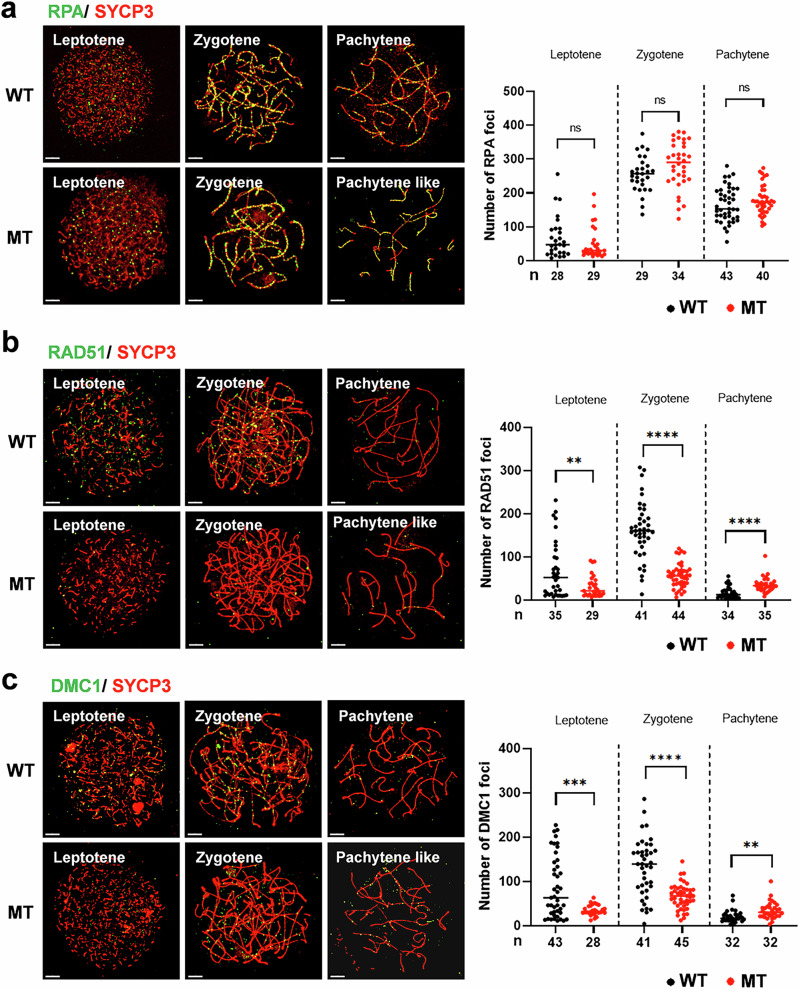
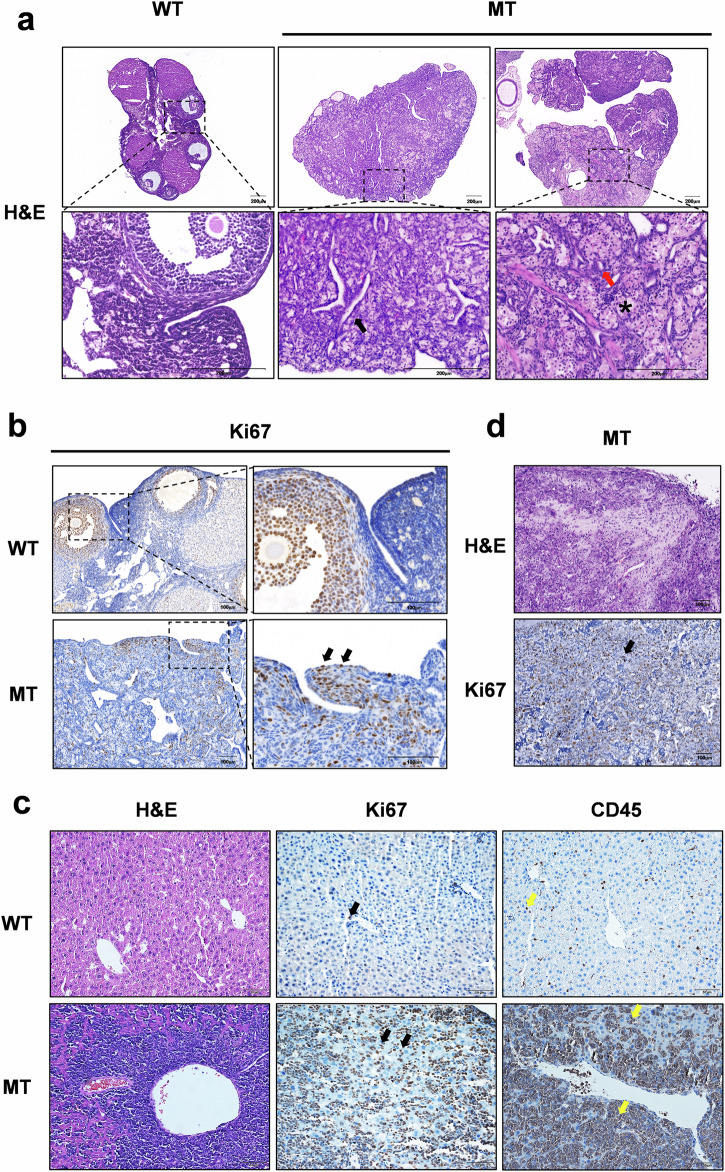
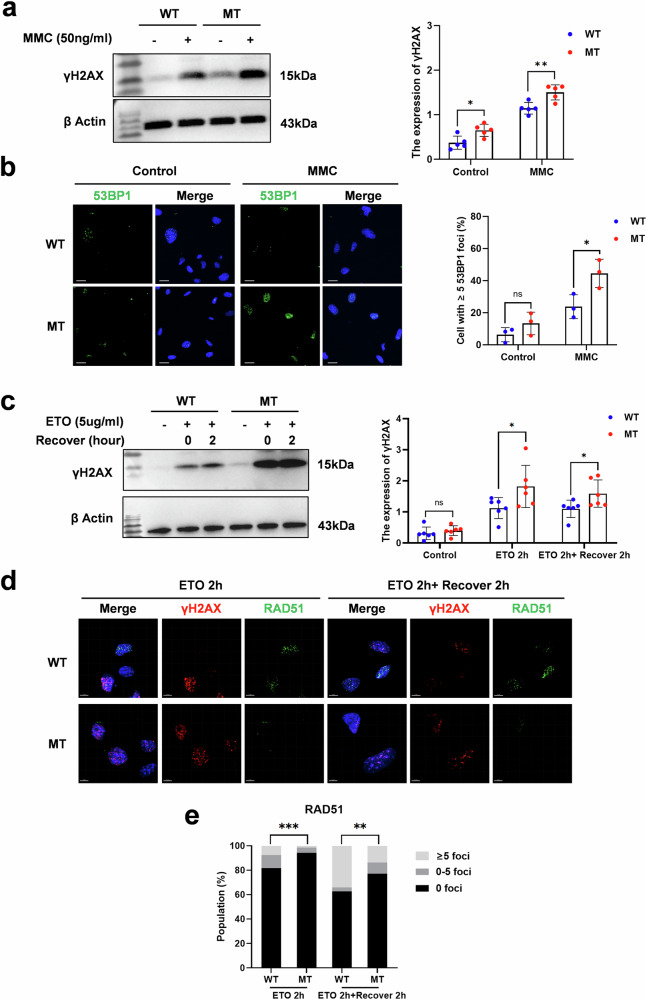
The DNA damage response plays a pivotal role in ovarian aging. Breast cancer susceptibility gene 2 (BRCA2), which participates in homologous recombination (HR), is a key regulator of natural menopause. Rare BRCA2 variants have been identified in patients with premature ovarian insufficiency (POI). However, the underlying molecular mechanisms are not well understood. Using a viable mouse model, Brca2c.68-1G>C/c.4384-4394del, carrying compound heterozygous variants mirroring the ones identified in a POI pedigree, we illustrated the essential role of BRCA2 in primordial follicle pool establishment. Germline deficiency of BRCA2 did not affect primordial germ cell (PGC) proliferation but impaired the recruitment of RAD51 and DMC1 to programmed DNA double-strand breaks (DSBs) during meiotic HR, causing postnatal oocyte depletion. Moreover, Brca2c.68-1G>C/c.4384-4394del mice presented increased tumor susceptibility. These findings confirmed the pathogenicity of BRCA2 biallelic variants in POI, revealing the dual impact on germ cell development and somatic cancer risk, underscoring the necessity of tumor surveillance in POI patients with BRCA2 mutations.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics statement: Ethical approval for the animal experiments was granted by the Institutional Review Board (IRB) of the Reproductive Medicine of Shandong University ([2014] IRB number: 52).
Figures
Similar articles
-
Distinct roles of the two BRCA2 DNA-binding domains in DNA damage repair and replication fork preservation.Cell Rep. 2025 May 27;44(5):115654. doi: 10.1016/j.celrep.2025.115654. Epub 2025 May 3. Cell Rep. 2025. PMID: 40323719
-
Rapamycin reduces DNA damage of in vitro matured oocytes by promoting Rad51-mediated homologous recombination.Reprod Biol Endocrinol. 2025 Jul 3;23(1):93. doi: 10.1186/s12958-025-01428-6. Reprod Biol Endocrinol. 2025. PMID: 40611176 Free PMC article.
-
BRCA Mutations, DNA Repair Deficiency, and Ovarian Aging.Biol Reprod. 2015 Sep;93(3):67. doi: 10.1095/biolreprod.115.132290. Epub 2015 Jul 29. Biol Reprod. 2015. PMID: 26224004 Free PMC article.
-
Methotrexate-mediated inhibition of RAD51 expression and homologous recombination in cancer cells.J Cancer Res Clin Oncol. 2012 May;138(5):811-8. doi: 10.1007/s00432-011-1132-8. Epub 2012 Jan 25. J Cancer Res Clin Oncol. 2012. PMID: 22274865 Free PMC article.
-
Positive and negative regulators of RAD51/DMC1 in homologous recombination and DNA replication.DNA Repair (Amst). 2024 Feb;134:103613. doi: 10.1016/j.dnarep.2023.103613. Epub 2023 Dec 13. DNA Repair (Amst). 2024. PMID: 38142595 Review.
References
-
- Baird, D. T. et al. Fertility and ageing. Hum. Reprod. Update11, 261–276 (2005). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
