

Repression via DNA looping by the Gram-positive global transcriptional regulator ScoC from Geobacillus
- PMID: 40715536
- PMCID: PMC12297555
- DOI: 10.1038/s42003-025-08555-1
Repression via DNA looping by the Gram-positive global transcriptional regulator ScoC from Geobacillus
Abstract
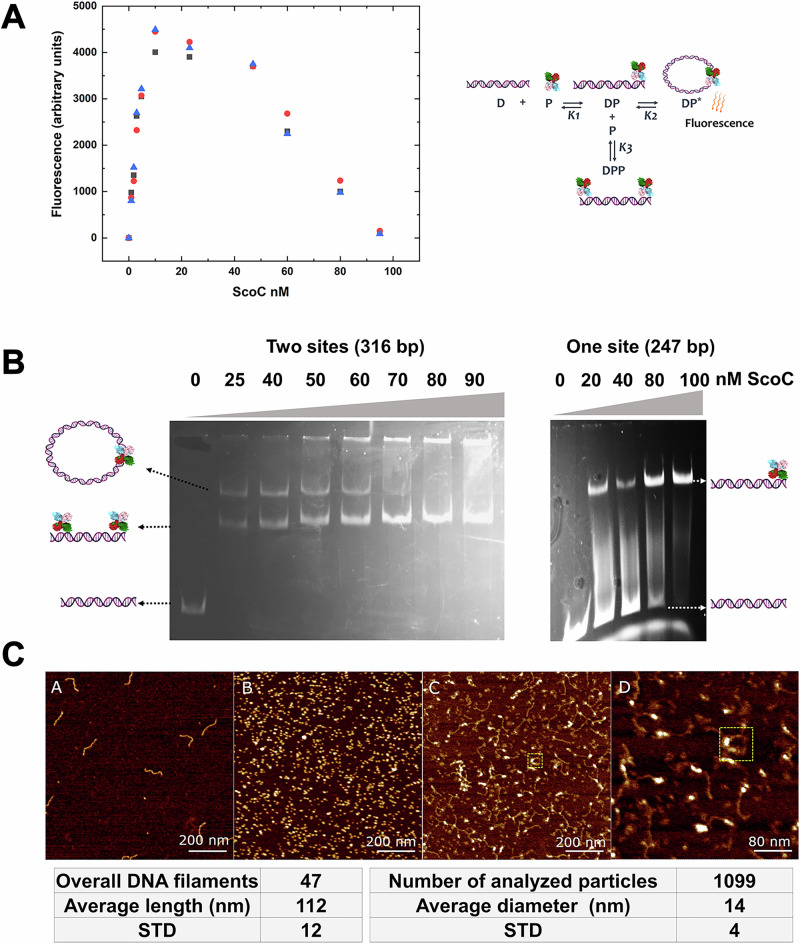
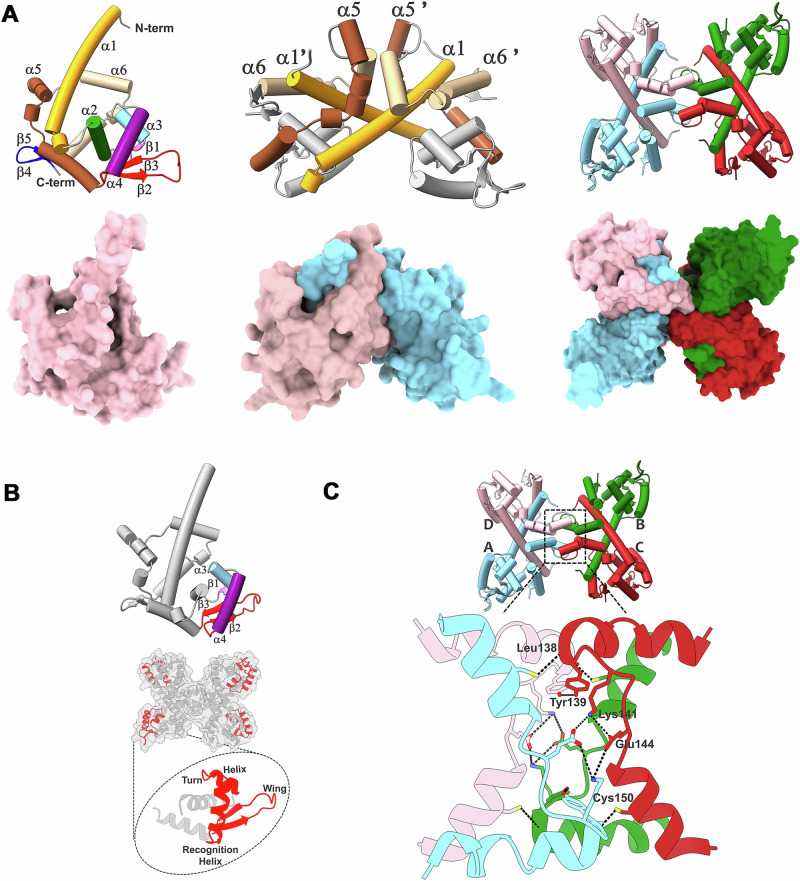
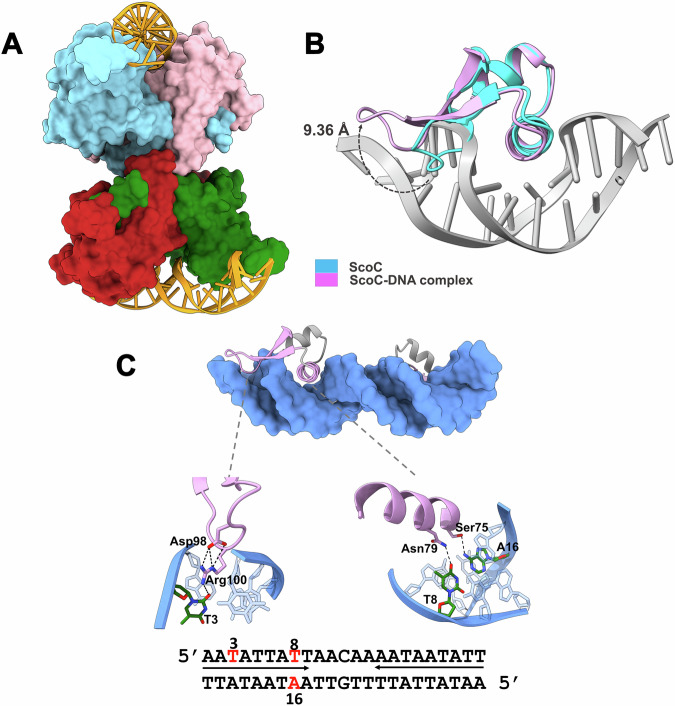
ScoC of the MarR family is a global regulator of transition phase pathways in Gram-positive bacteria, and in Bacillus subtilis it is estimated to regulate more than 500 genes. ScoC mediated activity is governed by its regulated expression, and by the interplay with other global transcriptional factors allowing for the finetuning of gene expression. Here we show, by transcriptional lacZ-fusions analysis, that ScoC from Geobacillus binds to two operator sites in the promoter region of the oligopeptide permease oppA, and that both binding sites are necessary for repression. Gel retardation assays, atomic force microscopy and fluorescence resonance energy transfer analyses demonstrate that ScoC can induce DNA looping. The crystal structures of ScoC and ScoC complexed with a 23-bp symmetric palindromic DNA from Geobacillus were determined at 3.15 Å and 3.50 Å resolution, respectively. The structures revealed a tetrameric X-shaped assembly composed of two dimers in which each dimeric unit comprises a winged helix-turn-helix DNA-binding motif. Our results expand the architecture of the MarR family regulators and suggest a mechanism by which ScoC interacts with other regulatory factors to modulate gene expression in the transition phase.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


Similar articles
-
Structural basis for higher-order DNA binding by a bacterial transcriptional regulator.PLoS Genet. 2025 Jun 27;21(6):e1011749. doi: 10.1371/journal.pgen.1011749. eCollection 2025 Jun. PLoS Genet. 2025. PMID: 40577318 Free PMC article.
-
RstA Is a Major Regulator of Clostridioides difficile Toxin Production and Motility.mBio. 2019 Mar 12;10(2):e01991-18. doi: 10.1128/mBio.01991-18. mBio. 2019. PMID: 30862746 Free PMC article.
-
Regulatory networks of FUR and NtcA are intertwined by transcriptional regulators, two-component systems, serine/threonine kinases, and sigma factors in Anabaena sp. PCC 7120.mSystems. 2025 Jul 22;10(7):e0037325. doi: 10.1128/msystems.00373-25. Epub 2025 Jun 25. mSystems. 2025. PMID: 40558093 Free PMC article.
-
Immunogenicity and seroefficacy of pneumococcal conjugate vaccines: a systematic review and network meta-analysis.Health Technol Assess. 2024 Jul;28(34):1-109. doi: 10.3310/YWHA3079. Health Technol Assess. 2024. PMID: 39046101 Free PMC article.
-
The effect of sample site and collection procedure on identification of SARS-CoV-2 infection.Cochrane Database Syst Rev. 2024 Dec 16;12(12):CD014780. doi: 10.1002/14651858.CD014780. Cochrane Database Syst Rev. 2024. PMID: 39679851 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
