

A designer polyQ fusion protein modulates NF-κB signaling by sequestering P65/RelA into aggregates
- PMID: 40717130
- PMCID: PMC12301440
- DOI: 10.1038/s41598-025-13237-3
A designer polyQ fusion protein modulates NF-κB signaling by sequestering P65/RelA into aggregates
Abstract
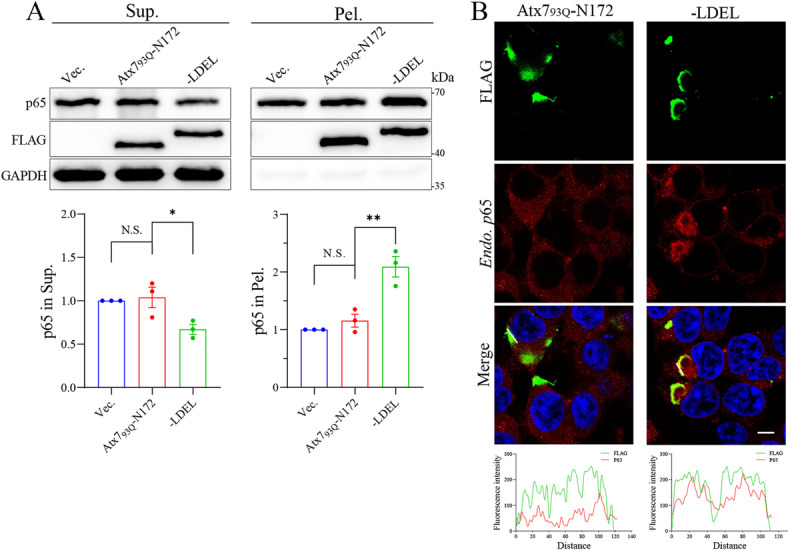
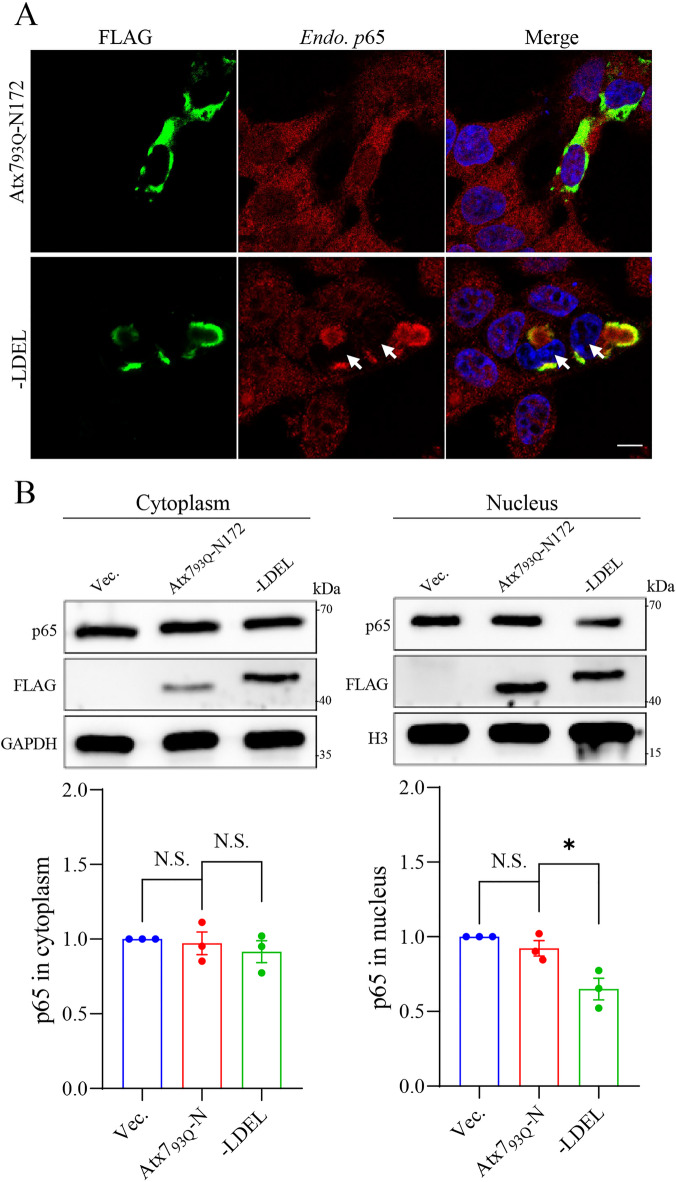
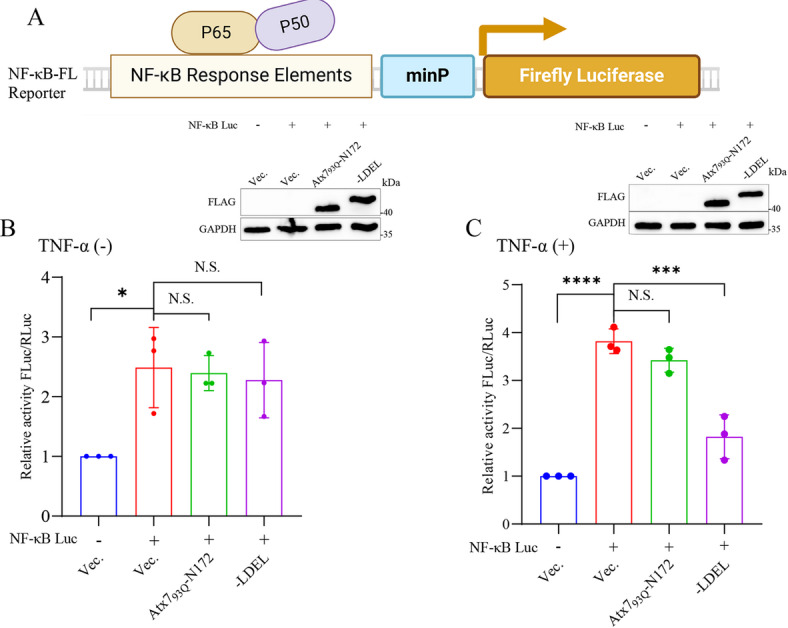
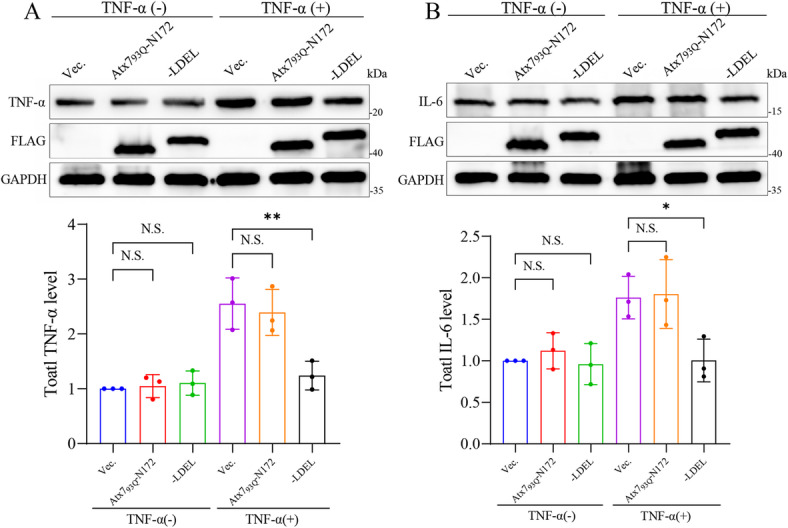
The transcription factor P65 is a subunit of the NF-κB complex that regulates expression of pro-inflammatory cytokines, thereby controlling the innate immune response. Excessive activation of NF-κB is commonly associated with various chronic inflammatory conditions. We previously established a polyglutamine (polyQ) fusion strategy to modulate biological processes by sequestering the targeted proteins. Here, we report a designer polyQ fusion (Atx793Q-N172-LDEL) that modulates the NF-κB signaling by sequestering P65 into aggregates. The fusion protein can interact with cellular P65 via its LDEL peptide sequence and specifically sequester it into aggregates or inclusions. This sequestration impedes the nuclear translocation process of P65, reduces its nuclear abundancy, and thereby attenuates the activity of NF-κB signaling and expression of the downstream genes, such as TNF-α and IL-6. This study provides therapeutic potential for treating inflammation and autoimmune disorders by targeting the P65 protein directly.
Keywords: Aggregation; LDEL peptide; NF-κB pathway.; P65 protein; PolyQ fusion; Sequestration.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

Similar articles
-
Buzhong Yiqi decoction improves inflammation and oxidative damage in autoimmune thyroiditis by inhibiting apoptosis via the SIRT1-Mediated Nrf2/NF-κB axis.J Ethnopharmacol. 2025 Jul 24;351:119967. doi: 10.1016/j.jep.2025.119967. Epub 2025 May 11. J Ethnopharmacol. 2025. PMID: 40360040
-
Biophysical characterization of RelA-p52 NF-κB dimer-A link between the canonical and the non-canonical NF-κB pathway.Protein Sci. 2024 Nov;33(11):e5184. doi: 10.1002/pro.5184. Protein Sci. 2024. PMID: 39412374
-
EVI1 acts as an inducible negative-feedback regulator of NF-κB by inhibiting p65 acetylation.J Immunol. 2012 Jun 15;188(12):6371-80. doi: 10.4049/jimmunol.1103527. Epub 2012 May 11. J Immunol. 2012. PMID: 22581859 Free PMC article.
-
The immunomodulatory effects of lactoferrin and its derived peptides on NF-κB signaling pathway: A systematic review and meta-analysis.Immun Inflamm Dis. 2023 Aug;11(8):e972. doi: 10.1002/iid3.972. Immun Inflamm Dis. 2023. PMID: 37647433 Free PMC article.
-
Neurotrophomodulatory effect of TNF-α through NF-κB in rat cortical astrocytes.Cytotechnology. 2025 Feb;77(1):37. doi: 10.1007/s10616-024-00698-z. Epub 2025 Jan 5. Cytotechnology. 2025. PMID: 39776978 Review.
References
-
- Housmans, J. A. J., Wu, G., Schymkowitz, J. & Rousseau, F. A guide to studying protein aggregation. FEBS J.290, 554–583. 10.1111/febs.16312 (2023). - PubMed
-
- Chiti, F. & Dobson, C. M. Protein misfolding amyloid formation, and human disease: a summary of progress over the last decade. Annu. Rev. Biochem.86, 27–68. 10.1146/annurev-biochem-061516-045115 (2017). - PubMed
-
- Arrasate, M., Mitra, S., Schweitzer, E. S., Segal, M. R. & Finkbeiner, S. Inclusion body formation reduces levels of mutant Huntingtin and the risk of neuronal death. Nature431, 805–810. 10.1038/nature02998 (2004). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
