

Metabolite-mediated interactions and direct contact between Fusobacterium varium and Faecalibacterium prausnitzii
- PMID: 40721806
- PMCID: PMC12302451
- DOI: 10.1186/s40168-025-02168-w
Metabolite-mediated interactions and direct contact between Fusobacterium varium and Faecalibacterium prausnitzii
Abstract
Background: The human gut harbors a diverse microbiota that is crucial for maintaining health but also contributes to several diseases. Understanding how microbial communities are assembled and maintained is critical for advancing gut health.
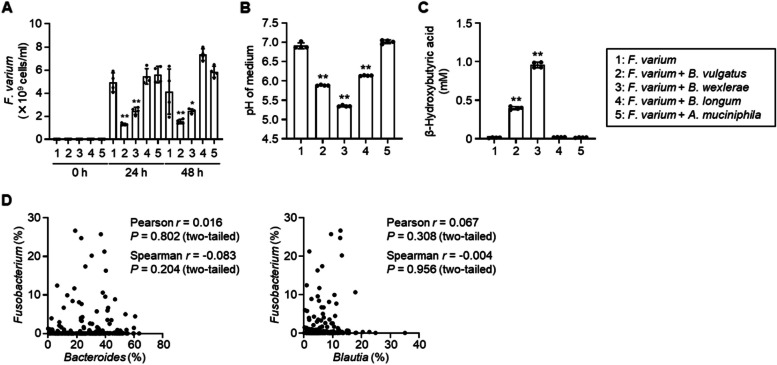
Results: We identified a unique interaction between the pathobiont Fusobacterium varium and the symbiont Faecalibacterium prausnitzii, both members of the gut microbial community; their interaction is driven by metabolites and direct cell-to-cell contact. Growth of F. varium was inhibited in the presence of F. prausnitzii because of a decrease in pH and an increase in β-hydroxybutyric acid. Conversely, the growth of F. prausnitzii was promoted in the presence of F. varium, likely via direct contact.
Conclusions: These findings highlight the importance of metabolite-driven interactions and direct contact in shaping gut microbial communities and emphasize the potential of interactions between F. prausnitzii and F. varium in influencing gut health. Video Abstract.
Keywords: Faecalibacterium; Fusobacterium; Gut ecosystem; Microbial interaction; Pathobiont; Symbiont.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: All experiments were approved by the Ethics Committee of the National Institutes of Biomedical Innovation, Health and Nutrition (NIBN) and were conducted in accordance with its guidelines (approval number: 296 m). Informed consent was obtained from all participants. Consent for publication: Not applicable. Competing interests: The authors have the following potential conflicts of interest: S. Maruyama, T. Matsuoka, and T. Kobayashi are employees of Hakubaku Co., Ltd. (Yamanashi, Japan). Other authors declare no competing interests.
Figures




References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
