

Membrane-Embedded Anti-Cancer Peptide Causes a Minimal Structural Perturbation That Is Sufficient to Enhance Phospholipid Flip-Flop and Charge Permeation Rates
- PMID: 40724510
- PMCID: PMC12300442
- DOI: 10.3390/life15071007
Membrane-Embedded Anti-Cancer Peptide Causes a Minimal Structural Perturbation That Is Sufficient to Enhance Phospholipid Flip-Flop and Charge Permeation Rates
Abstract
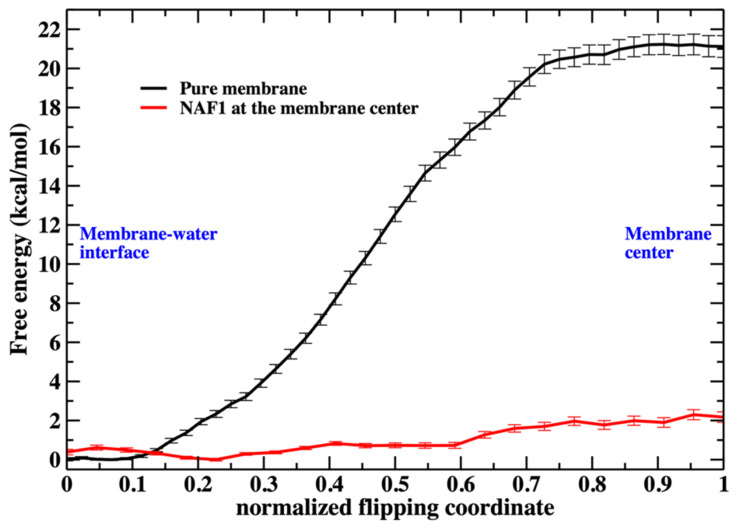
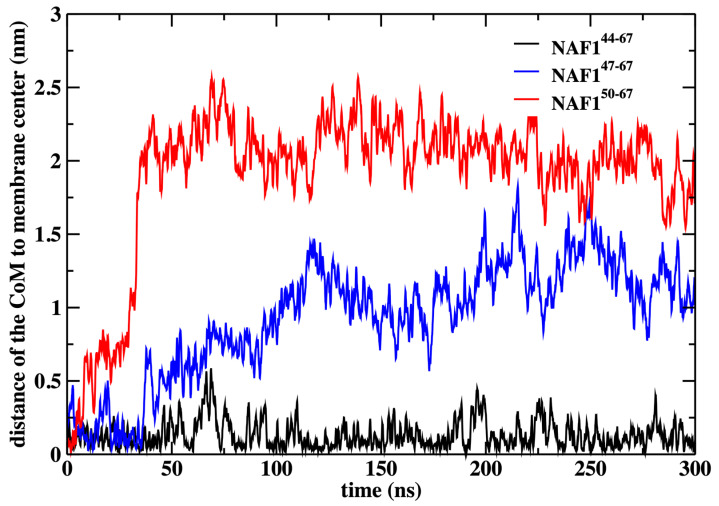
A prime role of biological membranes is to form barriers for material transport into and out of cells. Membranes consist of phospholipids with polar heads, which are presented to the aqueous solutions, and hydrophobic tails that form the membrane core. This construct prevents the permeation of hydrophilic, well-solvated molecules across the lipid hydrophobic barrier. The barrier is not absolute, and several approaches are available for efficient translocation. Channels and pumps enable selective and efficient transport across membranes. Another transport mechanism is passive permeation, in which permeants, without assistance, directly transport across membranes. Passive transport is coupled to transient defects in the membrane structure that make crossing the hydrophobic bilayer easier-for example, displacements of head groups from aqueous solution-membrane interface into the membrane core. The defects, in turn, are rare unless assisted by passively permeating molecules such as cell-penetrating peptides that distort the membrane structure. One possible defect is a phospholipid molecule with a head pointing to the hydrophobic core. This membrane distortion allows head group flipping from one layer to the other. We show computationally, using atomically detailed simulations and the Milestoning theory, that the presence of a cell-penetrating peptide in a membrane greatly increases phospholipid flip-flop rate and hence defect formation and the permeability of membranes.
Keywords: cell penetrating peptide; membrane defect; membrane permeation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures








Similar articles
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Comparison of cellulose, modified cellulose and synthetic membranes in the haemodialysis of patients with end-stage renal disease.Cochrane Database Syst Rev. 2001;(3):CD003234. doi: 10.1002/14651858.CD003234. Cochrane Database Syst Rev. 2001. Update in: Cochrane Database Syst Rev. 2005 Jul 20;(3):CD003234. doi: 10.1002/14651858.CD003234.pub2. PMID: 11687058 Updated.
-
Systemic Inflammatory Response Syndrome.2025 Jun 20. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jun 20. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31613449 Free Books & Documents.
-
Deep anterior lamellar keratoplasty versus penetrating keratoplasty for treating keratoconus.Cochrane Database Syst Rev. 2014 Jul 22;2014(7):CD009700. doi: 10.1002/14651858.CD009700.pub2. Cochrane Database Syst Rev. 2014. PMID: 25055058 Free PMC article.
-
Sexual Harassment and Prevention Training.2024 Mar 29. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Mar 29. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 36508513 Free Books & Documents.
References
-
- Sapay N., Bennett W.F.D., Tieleman D.P. Thermodynamics of flip-flop and desorption for a systematic series of phosphatidylcholine lipids. Soft Matter. 2009;5:3295–3302. doi: 10.1039/b902376c. - DOI
LinkOut - more resources
Full Text Sources
