

Neuroinflammation Based Neurodegenerative In Vitro Model of SH-SY5Y Cells-Differential Effects on Oxidative Stress and Insulin Resistance Relevant to Alzheimer's Pathology
- PMID: 40724831
- PMCID: PMC12294424
- DOI: 10.3390/ijms26146581
Neuroinflammation Based Neurodegenerative In Vitro Model of SH-SY5Y Cells-Differential Effects on Oxidative Stress and Insulin Resistance Relevant to Alzheimer's Pathology
Abstract
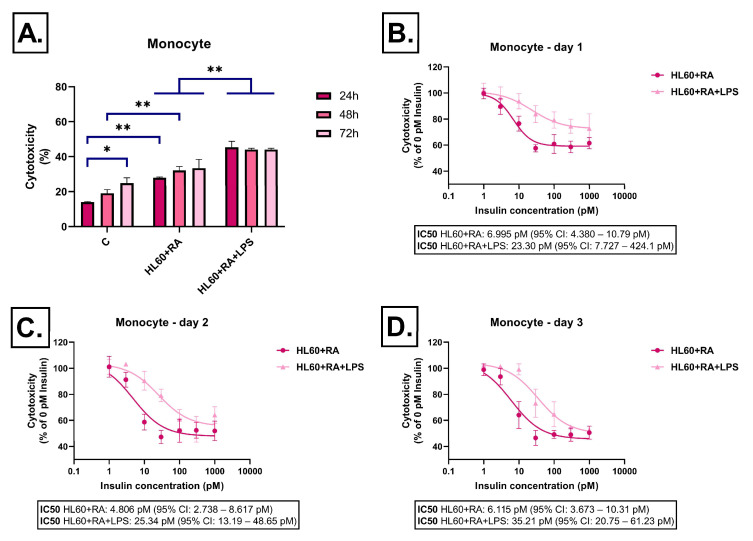
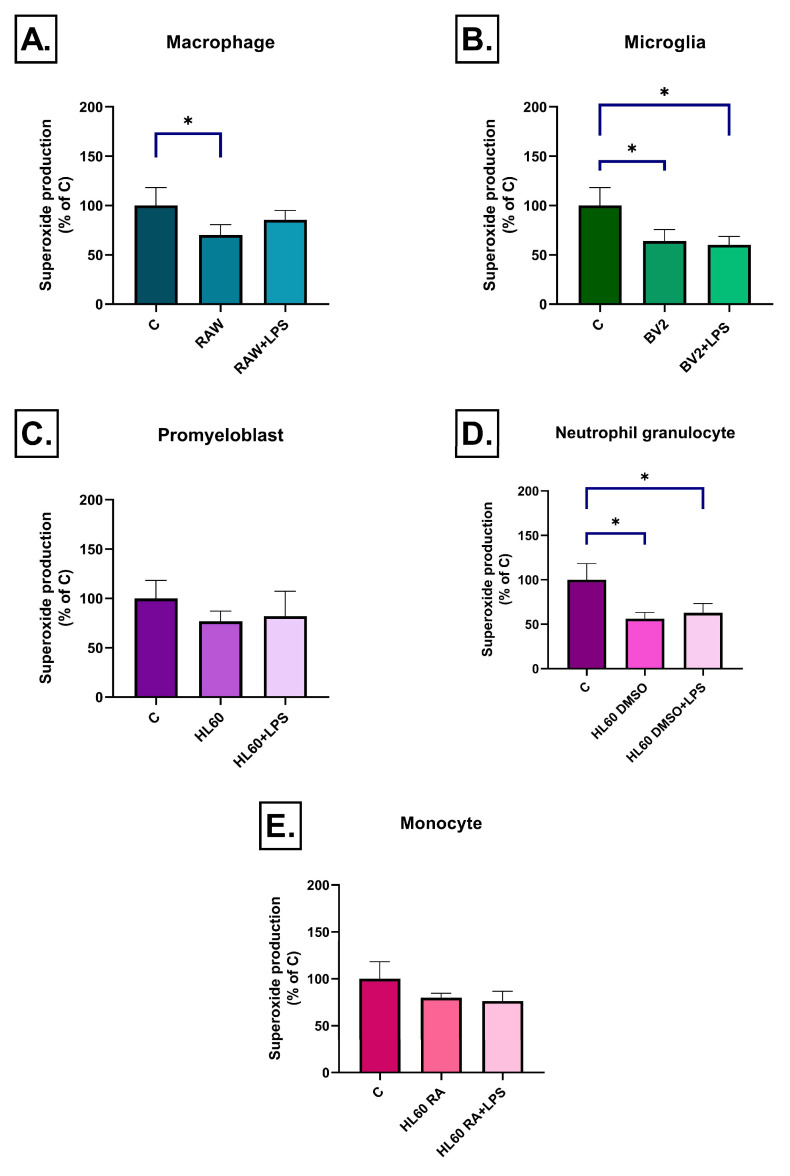
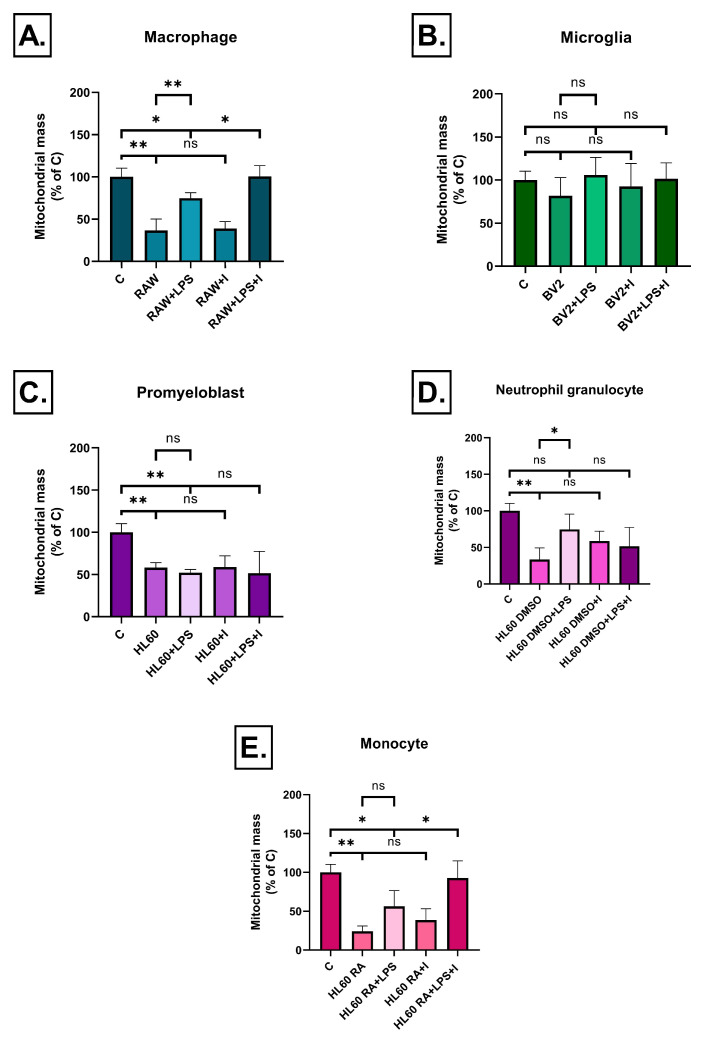
Neuroinflammation is a key process in Alzheimer's disease (AD). We aimed to examine the development and evaluation of a comprehensive in vitro model that captures the complex interplay between neurons and immune cell types. Retinoic acid-differentiated SH-SY5Y neuroblastoma cells exposed to LPS-conditioned media (CM) from RAW264.7 macrophages, BV2 microglia, and HL60 promyelocytic cells differentiated into neutrophil- or monocyte-like phenotypes were analyzed. The effects of CM containing inflammatory factors on neuronal viability and function were systematically evaluated. Neuronal oxidative stress, mitochondrial function, autophagy and protein aggregates were analyzed. The involvement of insulin resistance was studied by assaying glucose uptake and determining its IC50 values for cell viability improvement and GSK3β phosphorylation. After short-term exposure (3 h), most inflammatory CMs induced peroxide production in neurons, with the strongest effect observed in media from DMSO- or RA-differentiated HL60 cells. Mitochondrial membrane potential was markedly reduced by LPS-stimulated BV2 and HL60-derived CMs. Prolonged exposure (72 h) revealed partial normalization of oxidative stress and mitochondrial membrane potential. Glucose uptake was significantly impaired in cells treated with LPS-activated RAW264.7, BV2, and DMSO-differentiated HL60 cell media, while insulin partially rescued this effect, except for the CM of BV2 cells. Notably, insulin IC50 increased dramatically under LPS-treated BV2 cells induced inflammation (35 vs. 198 pM), confirming the development of insulin resistance. Immune cell-specific inflammation causes distinct effects on neuronal oxidative stress, mitochondrial function, protein aggregation, insulin signaling and viability. LPS-activated BV2-derived CM best recapitulates AD-related pathology, offering a relevant in vitro model for further studies.
Keywords: Alzheimer’s disease; insulin resistance; microglia; neuroinflammation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures















References
-
- Parbo P., Ismail R., Sommerauer M., Stokholm M.G., Hansen A.K., Hansen K.V., Amidi A., Schaldemose J.L., Gottrup H., Braendgaard H., et al. Does inflammation precede tau aggregation in early Alzheimer’s disease? A PET study. Neurobiol. Dis. 2018;117:211–216. doi: 10.1016/j.nbd.2018.06.004. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
