

Comparative Analysis of Diel and Circadian Eclosion Rhythms and Clock Gene Expression Between Sexes in the Migratory Moth Spodoptera frugiperda
- PMID: 40725335
- PMCID: PMC12296149
- DOI: 10.3390/insects16070705
Comparative Analysis of Diel and Circadian Eclosion Rhythms and Clock Gene Expression Between Sexes in the Migratory Moth Spodoptera frugiperda
Abstract
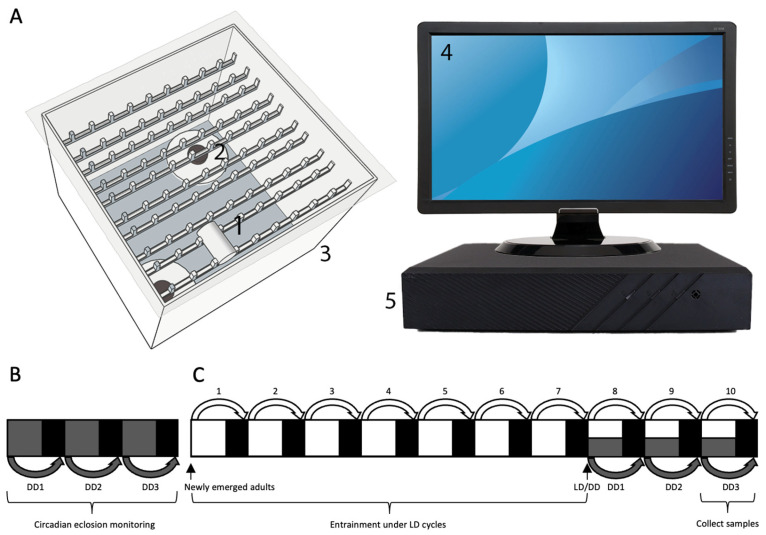
The circadian clock orchestrates behavioral and molecular processes such as eclosion. Understanding eclosion timing may offer insights into circadian mechanisms underlying migratory timing. Here, we characterize the diel and circadian patterns of eclosion and core clock gene expression in the fall armyworm (FAW), Spodoptera frugiperda, a globally distributed migratory moth. Using a custom-designed eclosion monitoring system under 14 h light: 10 h dark (L14: D10) and constant darkness (DD) conditions, we observed robust diel eclosion rhythms peaking shortly after lights-off under L14: D10, which became delayed and damped over three consecutive days in DD. Males showed a tendency toward more dispersed emergence patterns and exhibited statistically distinguishable eclosion distributions from females under both conditions. Expression of five canonical clock genes (cyc, clk, tim, per, cry2) displayed significant 24 h rhythmicity, with generally higher mesors in males. However, sex-specific differences in amplitude and phase were detected only for clk and cyc under L14: D10, not in DD. These findings suggest that sex-specific differences in circadian regulation are limited. Nonetheless, subtle variations in clock gene output and emergence timing in the FAW population established in China may contribute to sex-specific ecological strategies in the novel migratory arena.
Keywords: Spodoptera frugiperda; circadian rhythm; clock gene expression; eclosion timing; light regime; migratory insect pest; sex differences.
Conflict of interest statement
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures



References
-
- Duffy J.F., Cain S.W., Chang A.M., Phillips A.J., Munch M.Y., Gronfier C., Wyatt J.K., Dijk D.J., Wright K.P., Jr., Czeisler C.A. Sex difference in the near-24-hour intrinsic period of the human circadian timing system. Proc. Natl. Acad. Sci. USA. 2011;108:15602–15608. doi: 10.1073/pnas.1010666108. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
