

Systemic factors in young human serum influence in vitro responses of human skin and bone marrow-derived blood cells in a microphysiological co-culture system
- PMID: 40728515
- PMCID: PMC12339024
- DOI: 10.18632/aging.206288
Systemic factors in young human serum influence in vitro responses of human skin and bone marrow-derived blood cells in a microphysiological co-culture system
Abstract
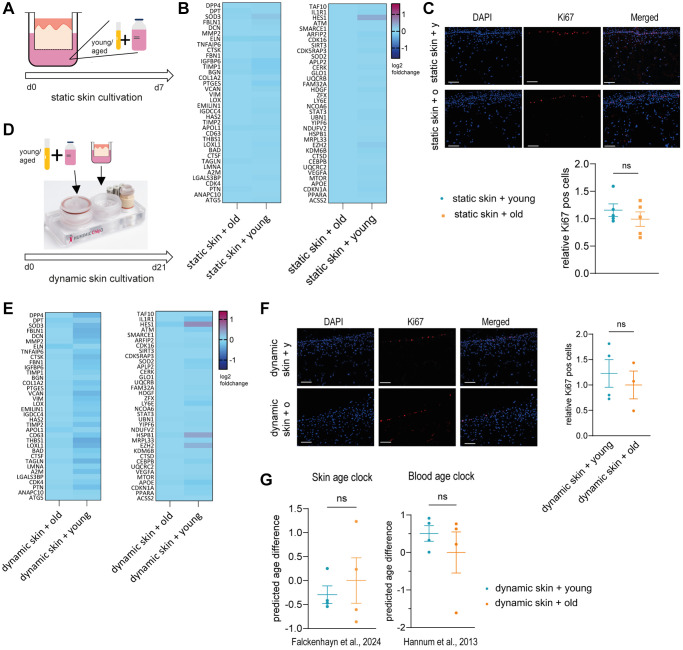
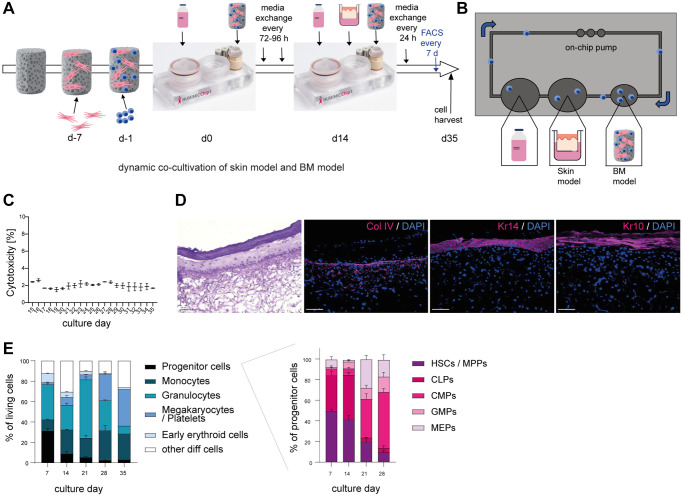
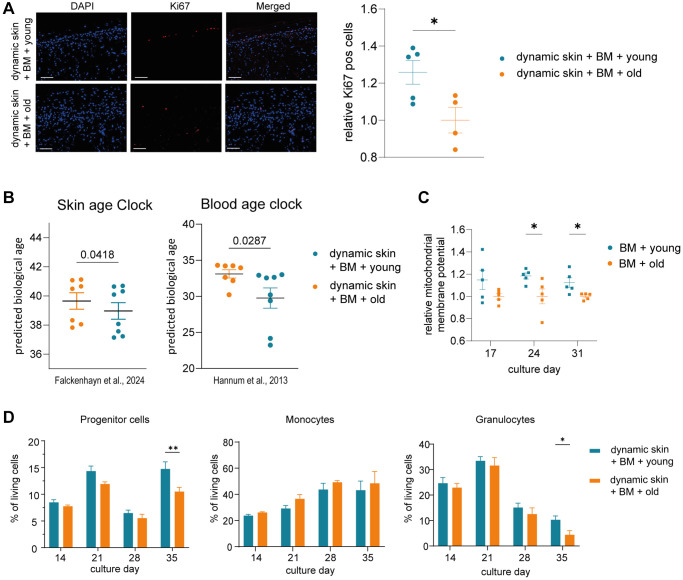
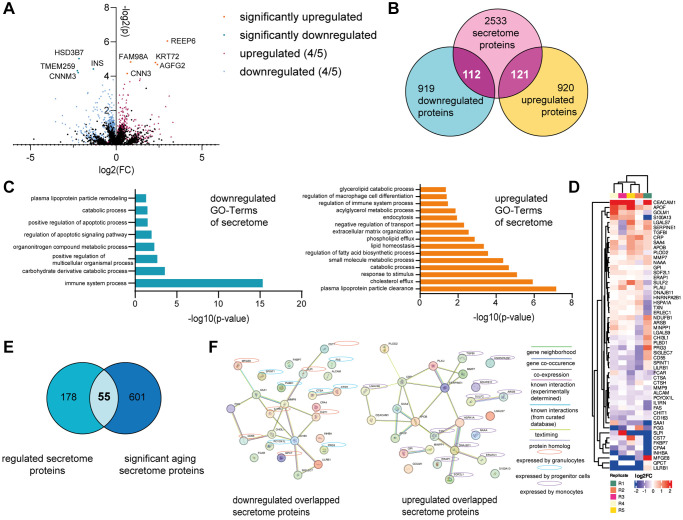
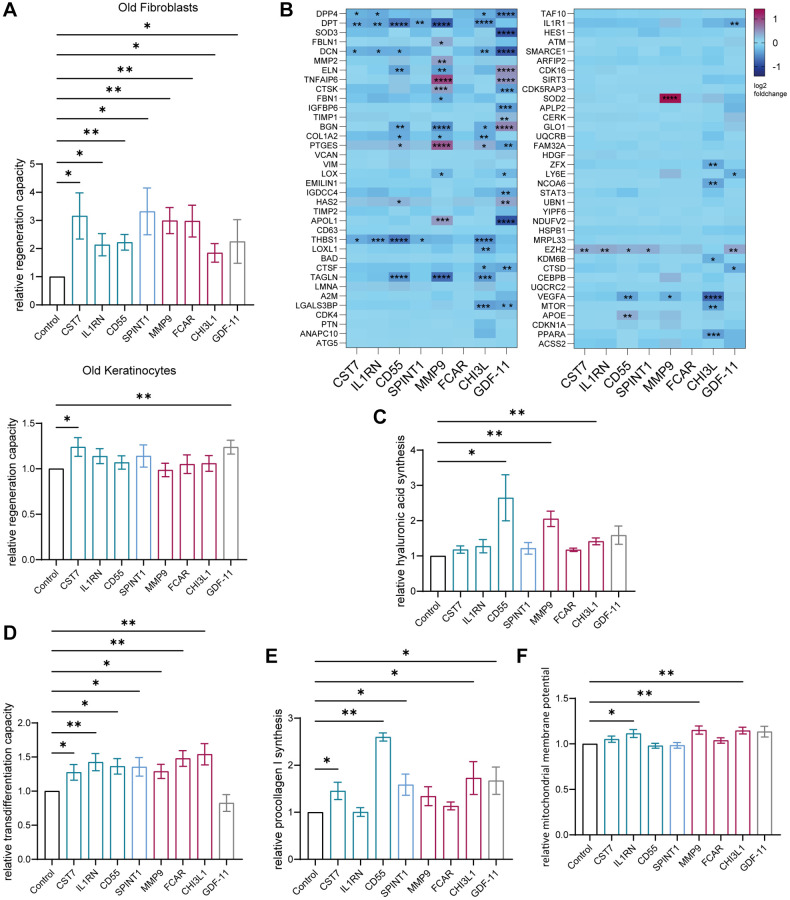
Aging is a complex process that significantly contributes to age-related diseases and poses significant challenges for effective interventions, with few holistic anti-aging approaches successfully reversing its signs. Heterochronic parabiosis studies illuminated the potential for rejuvenation through blood-borne factors, yet the specific drivers including underlying mechanisms remain largely unknown and until today insights have not been successfully translated to humans. In this study, we were able to recreate rejuvenation of the human skin via systemic factors using a microphysiological system including a 3D skin model and a 3D bone marrow model. Addition of young human serum in comparison to aged human serum resulted in an improvement of proliferation and a reduction of the biological age as measured by methylation-based age clocks in the skin tissue. Interestingly, this effect was only visible in the presence of bone marrow-derived cells. Further investigation of the bone marrow model revealed changes in the cell population in response to young versus aged human serum treatment. Using proteome analysis, we identified 55 potential systemic rejuvenating proteins produced by bone marrow-derived cells. For seven of these proteins, we were able to verify a rejuvenating effect on human skin cells using hallmarks of aging assays, supporting their role as systemic factors rejuvenating human skin tissue.
Keywords: bone marrow model; human serum; microphysiological systems; skin rejuvenation; systemic factors.
Conflict of interest statement
Figures
Similar articles
-
Prediction, screening and characterization of novel bioactive tetrapeptide matrikines for skin rejuvenation.Br J Dermatol. 2024 Jun 20;191(1):92-106. doi: 10.1093/bjd/ljae061. Br J Dermatol. 2024. PMID: 38375775
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2020 Jan 9;1(1):CD011535. doi: 10.1002/14651858.CD011535.pub3. Cochrane Database Syst Rev. 2020. Update in: Cochrane Database Syst Rev. 2021 Apr 19;4:CD011535. doi: 10.1002/14651858.CD011535.pub4. PMID: 31917873 Free PMC article. Updated.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
-
Rejuvenation of cerebromicrovascular function in aged mice through heterochronic parabiosis: insights into neurovascular coupling and the impact of young blood factors.Geroscience. 2024 Feb;46(1):327-347. doi: 10.1007/s11357-023-01039-2. Epub 2023 Dec 21. Geroscience. 2024. PMID: 38123890 Free PMC article.
References
-
- Pálovics R, Keller A, Schaum N, Tan W, Fehlmann T, Borja M, Kern F, Bonanno L, Calcuttawala K, Webber J, McGeever A, Luo J, Pisco AO, et al. , and Tabula Muris Consortium. Molecular hallmarks of heterochronic parabiosis at single-cell resolution. Nature. 2022; 603:309–14. 10.1038/s41586-022-04461-2 - DOI - PMC - PubMed
-
- Horvath S, Singh K, Raj K, Khairnar SI, Sanghavi A, Shrivastava A, Zoller JA, Li CZ, Herenu CB, Canatelli-Mallat M, Lehmann M, Habazin S, Novokmet M, et al. Reversal of biological age in multiple rat organs by young porcine plasma fraction. Geroscience. 2024; 46:367–94. 10.1007/s11357-023-00980-6 - DOI - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
