

Experience-Dependent Intrinsic Plasticity in Layer IV of Barrel Cortex at Whisking Onset
- PMID: 40730470
- PMCID: PMC12364433
- DOI: 10.1523/ENEURO.0252-25.2025
Experience-Dependent Intrinsic Plasticity in Layer IV of Barrel Cortex at Whisking Onset
Abstract
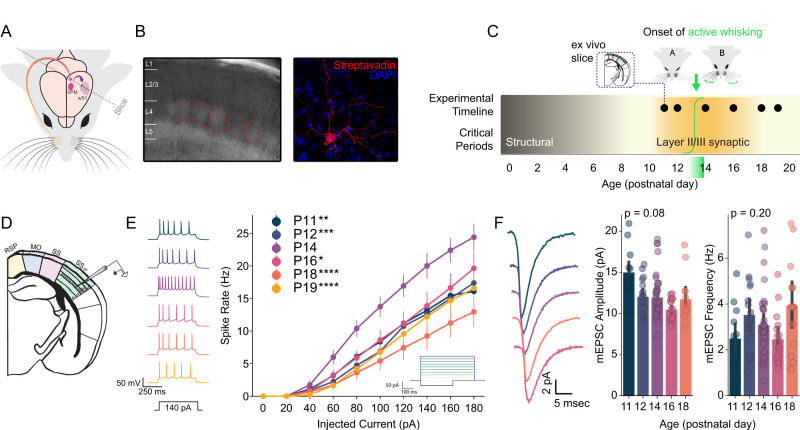
The development of motor control over sensory organs is a critical milestone, enabling active exploration and shaping of the sensory environment. Whether the onset of sensory organ motor control directly influences the development of corresponding sensory cortices remains unknown. Here, we confirm and exploit the late onset of whisking behavior in mice to address this question in the somatosensory system. Using ex vivo electrophysiology, we describe a transient increase in the intrinsic excitability of excitatory neurons in layer IV of the barrel cortex, which processes whisker input, immediately following the onset of active whisking on postnatal days 13 and 14. This increase in neuronal gain is specific to layer IV, independent of changes in synaptic strength, and requires prior sensory experience. Further, these effects are not expressed in inhibitory interneurons in barrel cortex. The transient increase in excitability is not evident in layer II/III of barrel cortex or in the visual cortex upon eye opening, suggesting a unique interaction between the development of active sensing and the thalamocortical input layer in the somatosensory isocortex. Predictive modeling indicates that, immediately following the onset of active whisking, changes in active membrane conductances alone can reliably distinguish neurons in control but not whisker-deprived hemispheres. Our findings demonstrate an experience-dependent, lamina-specific refinement of neuronal excitability tightly linked to the emergence of active whisking. This transient increase in the gain of the thalamic input layer coincides with a critical period for synaptic plasticity in downstream layers, suggesting a role in cortical maturation and sensory processing.
Keywords: barrel cortex; behavior; development; excitability, plasticity.
Copyright © 2025 Shallow et al.
Conflict of interest statement
The authors declare no competing financial interests.
Figures





Similar articles
-
Immediately following the onset of active whisking, the input layer of barrel cortex exhibits a transient period of increased excitability that depends on prior experience.bioRxiv [Preprint]. 2025 Apr 25:2024.06.04.597353. doi: 10.1101/2024.06.04.597353. bioRxiv. 2025. PMID: 38895408 Free PMC article. Preprint.
-
Peripuberty Is a Sensitive Period for Prefrontal Parvalbumin Interneuron Activity to Impact Adult Cognitive Flexibility.Dev Neurosci. 2025;47(2):127-138. doi: 10.1159/000539584. Epub 2024 Jun 3. Dev Neurosci. 2025. PMID: 38830346 Free PMC article.
-
Impairment in the homeostatic recruitment of layer 5/6 neurons following whisker stimulation in Fmr1 KO mice.Neurobiol Dis. 2025 Apr;207:106837. doi: 10.1016/j.nbd.2025.106837. Epub 2025 Feb 10. Neurobiol Dis. 2025. PMID: 39938578 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Bioengineered nerve conduits and wraps for peripheral nerve repair of the upper limb.Cochrane Database Syst Rev. 2022 Dec 7;12(12):CD012574. doi: 10.1002/14651858.CD012574.pub2. Cochrane Database Syst Rev. 2022. PMID: 36477774 Free PMC article.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources