

Variations in flanking or less conserved positions of Reb1 and Abf1 consensus binding sites lead to major changes in their ability to modulate nucleosome sliding activity
- PMID: 40731031
- PMCID: PMC12305957
- DOI: 10.1186/s40659-025-00627-0
Variations in flanking or less conserved positions of Reb1 and Abf1 consensus binding sites lead to major changes in their ability to modulate nucleosome sliding activity
Abstract
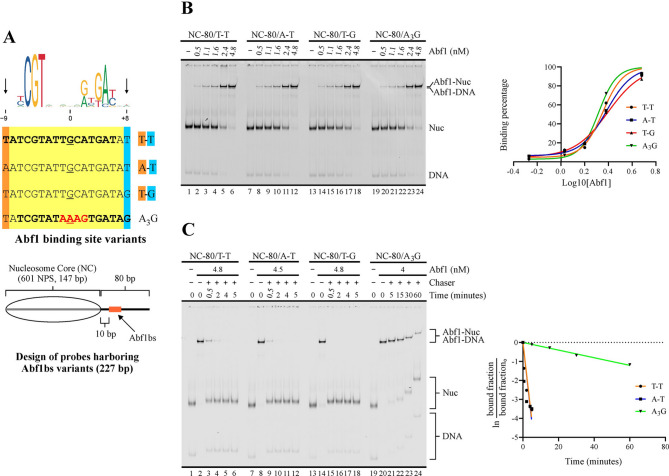
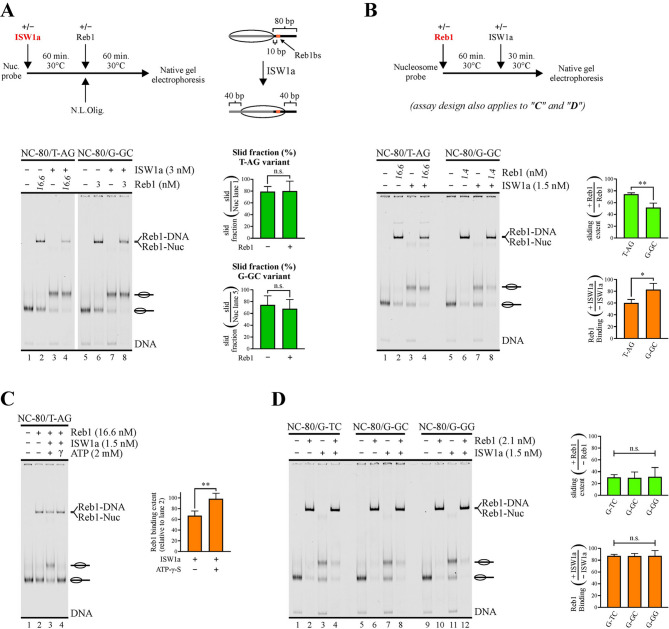
Background: Maintenance of nucleosome-free regions at gene regulatory regions conform a relevant aspect within chromatin dynamics. In the yeast Saccharomyces cerevisiae, Reb1 and Abf1 are among the transcriptions factors that perform this molecular function. These factors are thought to act as a barrier to nucleosome sliding that chromatin remodeling complexes such as ISW1a perform towards this region, being binding affinity a critical feature to act as a barrier. In this regard, sequence variations at positions flanking transcription factor binding sites could affect DNA shape features and, in turn, binding strength. In addition, recent studies have shown that positions of low conservation and/or flanking sequences might vary from gene bodies to gene regulatory regions. Considering these issues, we aimed to analyze whether variations in flanking or less conserved positions of Reb1 and Abf1 target sequences affect their binding affinity, especially dwell time, and their ability to hinder ISW1a's sliding activity.
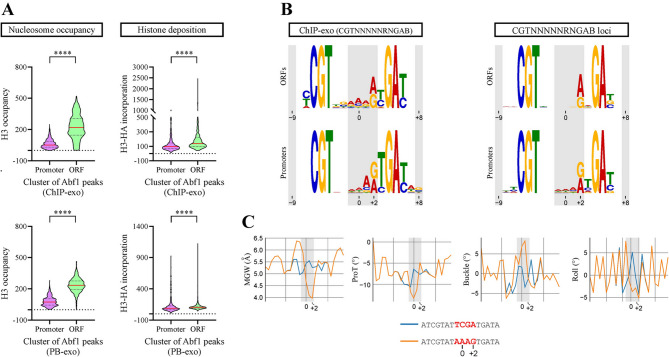
Results: We found that sequence changes at these positions deeply affect binding strength, particularly dwell time, and the ability to hinder ISW1a's sliding activity. Importantly, even under conditions where a markedly higher transcription factor concentration for a weak binding site was used to compare it to a strong binding site under an equal binding saturation level, the strong site displayed a significantly higher ability to hinder sliding activity. Moreover, genome-wide analyses showed that the sequence variants of Reb1 and Abf1 binding sites conferring this ability to hinder sliding activity to these factors are enriched at promoter regions relative to gene bodies.
Conclusions: Our findings show that dwell time is a key feature to hinder nucleosome sliding activity. For Reb1 and Abf1 factors, sequence variation at less conserved positions of their binding sites strongly affects this feature. The differential frequency at these positions found at promoter regions, relative to gene bodies, highlights the relevance of including this type of comparison in certain strategies used to determine the consensus binding site for transcription factors. To determine the molecular functions that require long dwell times and the transcription factors responsible for these tasks will significantly contribute to untangle the grammar of cis-regulatory elements.
Keywords: Abf1; Chromatin dynamics; Chromatin remodeling; Dwell time; ISW1a; Nucleosome remodeling; Nucleosome sliding; Reb1; Residence time; TFBS motifs.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: Not applicable. Competing interests: The authors declare that they have no competing interests.
Figures



Similar articles
-
General regulatory factors exert differential effects on nucleosome sliding activity of the ISW1a complex.Biol Res. 2024 May 4;57(1):22. doi: 10.1186/s40659-024-00500-6. Biol Res. 2024. PMID: 38704609 Free PMC article.
-
Home treatment for mental health problems: a systematic review.Health Technol Assess. 2001;5(15):1-139. doi: 10.3310/hta5150. Health Technol Assess. 2001. PMID: 11532236
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
References
-
- Lieleg C, Krietenstein N, Walker M, Korber P. Nucleosome positioning in yeasts: methods, maps, and mechanisms. Chromosoma. 2015;124(2):131–51. - PubMed
-
- Kubik S, Bruzzone MJ, Challal D, Dreos R, Mattarocci S, Bucher P, et al. Opposing chromatin remodelers control transcription initiation frequency and start site selection. Nat Struct Mol Biol. 2019;26(8):744–54. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
