

Unleash the dogs of death: metacaspase 5, microtubules, and hypersensitive response
- PMID: 40736719
- PMCID: PMC12310787
- DOI: 10.1007/s00299-025-03567-x
Unleash the dogs of death: metacaspase 5, microtubules, and hypersensitive response
Abstract
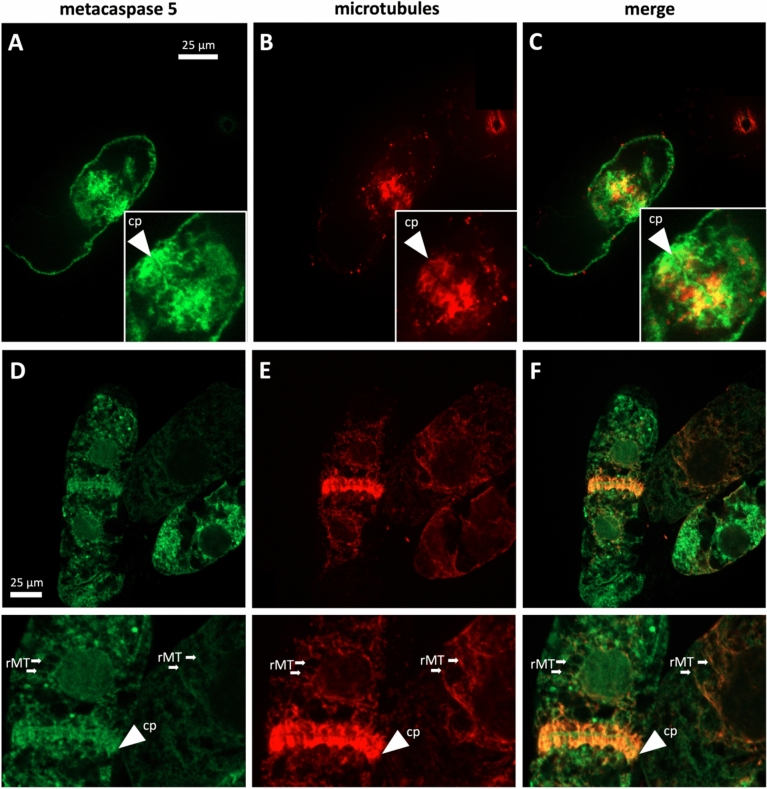
Vitis rupestris metacaspase 5, tethered to microtubules, drives grapevine. Hypersensitive response via calcium-dependent auto-processing, linking cytoskeletal dynamics to defence activation by elicitors. Metacaspase 5 is a key player for the hypersensitive response of grapevine against biotrophic pathogens and must be activated rapidly as to prevent colonisation. This activation is likely to occur through changes in protein activity. By expressing a GFP fusion of metacaspase 5 from Vitis rupestris in tobacco BY-2 cells, we can show that this protein is bound to microtubules and that the overexpressors are more responsive to the cell-death-inducing elicitors, cis-3-hexenal and harpin. The disruption of microtubules and actin filaments by these elicitors can be blocked by inhibitors of dynamic turnover and stabilisation. Stabilisation of microtubules by taxol can mitigate cis-3-hexenal induced mortality. Mutations of the catalytic or the putative microtubule-binding sites of metacaspase 5 can suppress auto-processing of this enzyme in biochemical assay. Likewise, the response to cis-3-hexenal (cell death, induction of salicylate-related gene expression) is suppressed in cells, whilst the cytoplasmic remodelling is retained. Calcium and the sites for catalysis or microtubule binding are required for both auto-processing and enzyme activity. We arrive at a model, where metacaspase 5 is inactive when tethered to microtubules, but becomes unleashed for auto-processing upon defence-mediated microtubule breakdown.
Keywords: Cis-3-hexenal; Vitis rupestris; Hypersensitive response; Metacaspase; Programmed cell death; Tobacco BY-2.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Conflict of interest: The authors declare they do not have any conflict of interest whatsoever.
Figures









References
-
- Brooks DM, Bender CL, Kunkel BN (2005) The Pseudomonas syringae phytotoxin coronatine promotes virulence by overcoming salicylic acid-dependent defences in A. thaliana. Mol Plant Pathol 6:629–639 - PubMed
-
- Buschmann H, Green P, Sambade A, Doonan JH, Lloyd CW (2011) Cytoskeletal dynamics in interphase, mitosis and cytokinesis analysed through Agrobacterium mediated transient transformation of tobacco BY-2 cells. New Phytol 190:258–267 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
