

Direct interaction of HMGB1 with SARS-CoV-2 facilitates its infection via RAGE-dependent endocytosis
- PMID: 40740486
- PMCID: PMC12307675
- DOI: 10.1016/j.isci.2025.113063
Direct interaction of HMGB1 with SARS-CoV-2 facilitates its infection via RAGE-dependent endocytosis
Abstract
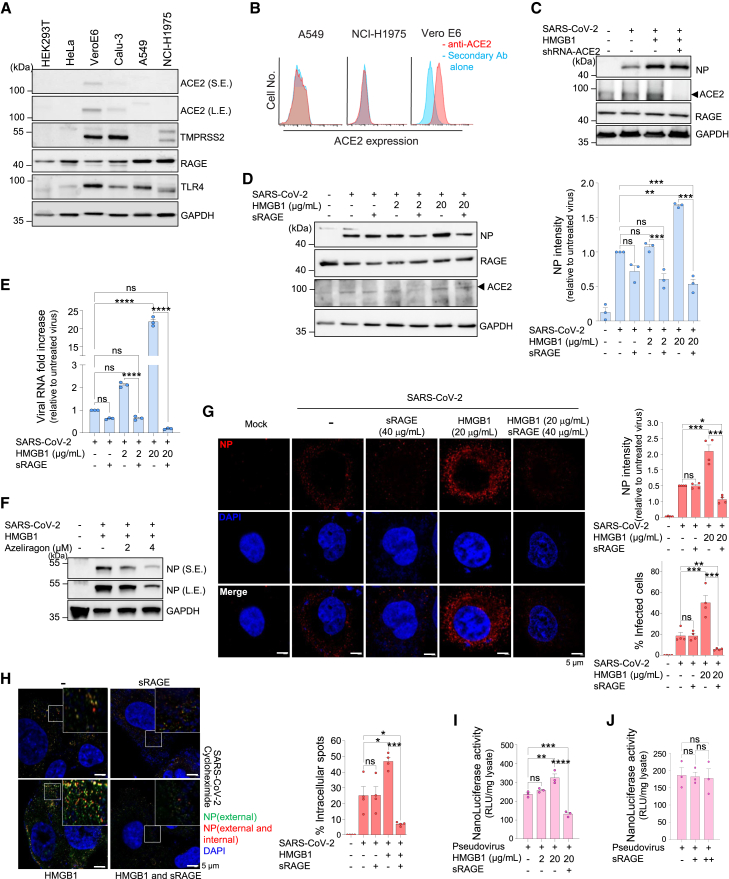
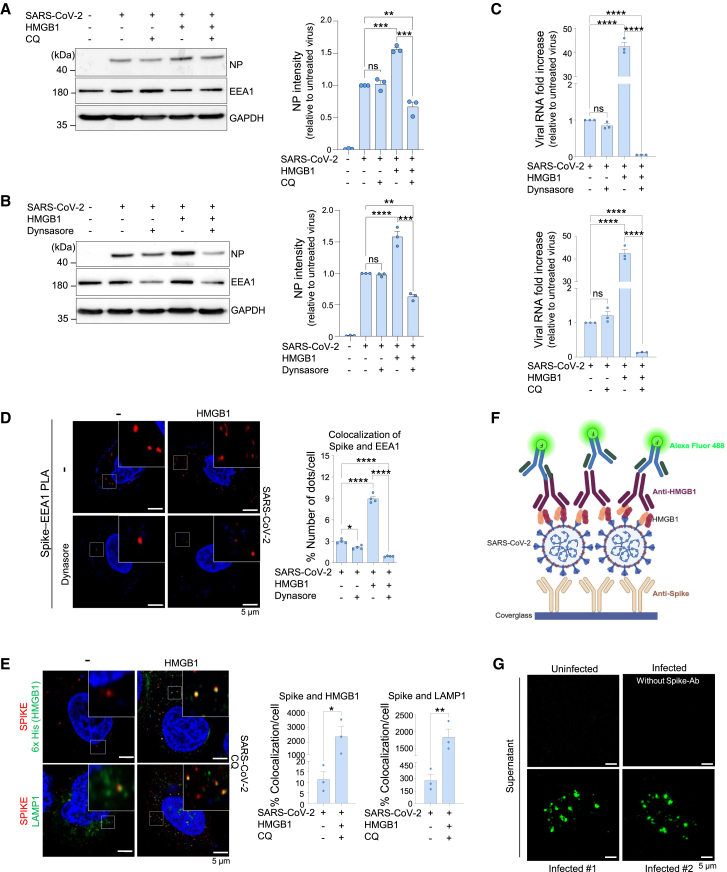
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the causative agent of coronavirus disease 2019 (COVID-19), has caused >770 million infections since 2020. SARS-CoV-2 outcomes are largely influenced by host immune responses. Among key mediators of innate immunity, high-mobility group box 1 (HMGB1) has gained attention for its role in inflammation during SARS-CoV-2 infection, and its levels are significantly elevated in acute and post-COVID-19 cases, correlating with disease severity. This study investigated the role of HMGB1 in COVID-19 pathogenesis. Our findings demonstrate that the SARS-CoV-2 spike protein directly interacts with HMGB1, forming an HMGB1-SARS-CoV-2 complex. This complex interacts with the receptor for advanced glycation end-products (RAGE), facilitating clathrin-mediated endocytosis and enhancing SARS-CoV-2 infection in human lung cells in vitro and in mouse models of infection. Overall, this study demonstrates the role of HMGB1 in promoting viral entry via RAGE, emphasizing its potential as a therapeutic target in severe COVID-19 cases.
Keywords: Cell biology; Immunology; Virology.
© 2025 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- World Health Organization 2023 data.who.int, WHO Coronavirus (COVID-19) dashboard > More resources [Dashboard] https://data.who.int/dashboards/covid19/more-resources
-
- Arunachalam P.S., Wimmers F., Mok C.K.P., Perera R.A.P.M., Scott M., Hagan T., Sigal N., Feng Y., Bristow L., Tak-Yin Tsang O., et al. Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science. 2020;369:1210–1220. doi: 10.1126/science.abc6261. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
