

Global divergence in plant and mycorrhizal fungal diversity hotspots
- PMID: 40744917
- PMCID: PMC12314030
- DOI: 10.1038/s41467-025-60106-8
Global divergence in plant and mycorrhizal fungal diversity hotspots
Abstract
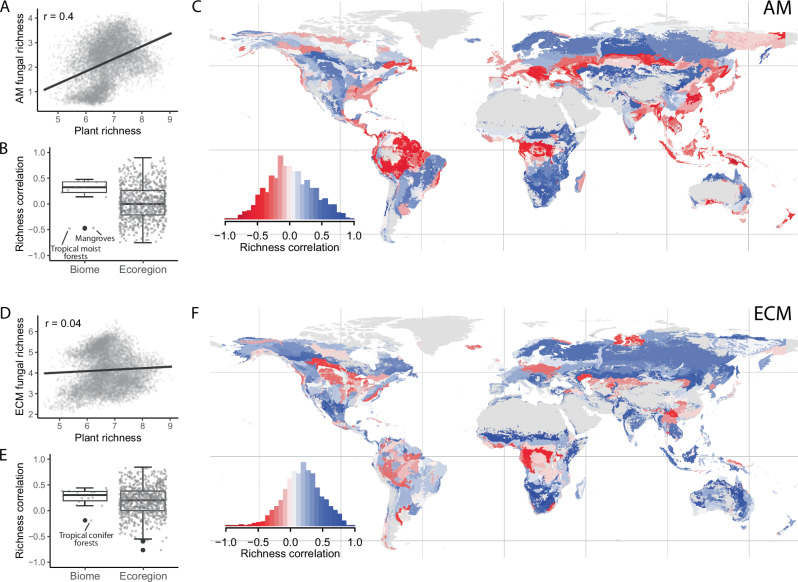
Environmental protection strategies often rely on aboveground biodiversity indicators for prioritising conservation efforts. However, substantial biodiversity exists belowground, and it remains unclear whether aboveground diversity hotspots are indicative of high soil biodiversity. Using geospatial layers of vascular plant, arbuscular mycorrhizal fungi, and ectomycorrhizal fungi alpha diversity, we map plant-fungal diversity associations across different scales and evaluate evidence for potential correlation drivers. Plant-fungal diversity correlations are weak at the global scale but stronger at regional scales. Plant-arbuscular mycorrhizal fungal correlations are generally negative in forest biomes and positive in grassland biomes, whereas plant-ectomycorrhizal fungal correlations are mostly positive or neutral. We find evidence that symbiosis strength, environmental covariation, and legacy effects all influence correlation patterns. Only 8.8% of arbuscular mycorrhizal and 1.5% of ectomycorrhizal fungal diversity hotspots overlap with plant hotspots, indicating that prioritising conservation based solely on aboveground diversity may fail to capture diverse belowground regions.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



Similar articles
-
Diversity and spatial structure of belowground plant-fungal symbiosis in a mixed subtropical forest of ectomycorrhizal and arbuscular mycorrhizal plants.PLoS One. 2014 Jan 28;9(1):e86566. doi: 10.1371/journal.pone.0086566. eCollection 2014. PLoS One. 2014. PMID: 24489745 Free PMC article.
-
Mycorrhiza increases plant diversity and soil carbon storage in grasslands.Proc Natl Acad Sci U S A. 2025 Feb 18;122(7):e2412556122. doi: 10.1073/pnas.2412556122. Epub 2025 Feb 12. Proc Natl Acad Sci U S A. 2025. PMID: 39937867 Free PMC article.
-
The impact of land use change on mycorrhizal fungi and their associations with rodents: insights from a temperate forest in Mexico.Mycorrhiza. 2025 May 8;35(3):36. doi: 10.1007/s00572-025-01210-x. Mycorrhiza. 2025. PMID: 40338382 Free PMC article.
-
Advancing knowledge on the biogeography of arbuscular mycorrhizal fungi to support Sustainable Development Goal 15: Life on Land.FEMS Microbiol Lett. 2025 Jan 10;372:fnaf055. doi: 10.1093/femsle/fnaf055. FEMS Microbiol Lett. 2025. PMID: 40539942 Free PMC article. Review.
-
Coupling of plant and mycorrhizal fungal diversity: its occurrence, relevance, and possible implications under global change.New Phytol. 2022 Jun;234(6):1960-1966. doi: 10.1111/nph.17954. Epub 2022 Jan 29. New Phytol. 2022. PMID: 35014033 Review.
References
-
- Tilman, D., Isbell, F. & Cowles, J. M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst.45, 471–493 (2014).
-
- Soliveres, S. et al. Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality. Nature536, 456–459 (2016). - PubMed
-
- Bardgett, R. D. & van der Putten, W. H. Belowground biodiversity and ecosystem functioning. Nature515, 505–511 (2014). - PubMed
-
- Van Der Heijden, M. G. A., Bardgett, R. D. & Van Straalen, N. M. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett.11, 296–310 (2008). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
