

Exogenous Nucleotides Supplementation Attenuates Age-Related Sarcopenia
- PMID: 40745399
- PMCID: PMC12313545
- DOI: 10.1002/jcsm.70002
Exogenous Nucleotides Supplementation Attenuates Age-Related Sarcopenia
Abstract
Background: Sarcopenia, an age-related clinical syndrome characterized by reduced skeletal muscle mass and strength, often leads to a loss of physical function. Nucleotides (NTs) supplementation is a potential strategy for preventing age-related sarcopenia as evidence suggests that NTs declined in muscles with aging, and 5'-CMP, 5'-UMP can mitigate muscle atrophy in C2C12 myotubes. This study aimed to investigate the effects of NTs supplementation on sarcopenia and its possible mechanism.
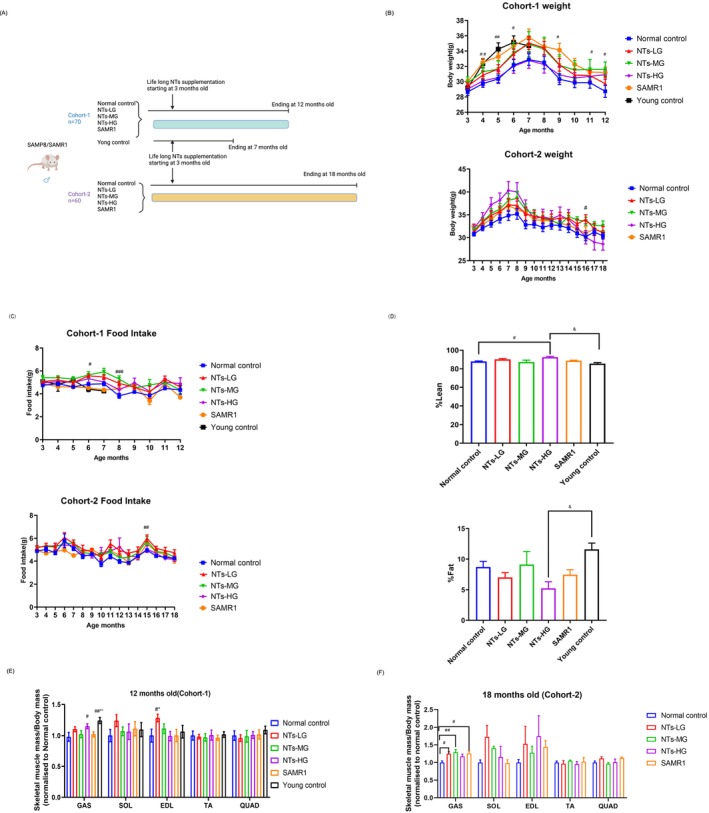
Methods: Senescence-accelerated mouse prone-8 (SAMP8) mice and C2C12 cells were used to assess the effect of NTs on sarcopenia. We fed an NTs-enriched mixture to SAMP8 mice starting from the age of 3 months for 9 months or 15 months. NTs' effects on H2O2-induced C2C12 atrophy cells were tested using nucleotide monomers and mixtures. The body composition was measured by EchoMRI analysis meter. Physical performance was tested including grip strength, wire hang, horizontal bar and gait test. The immunofluorescence staining was used to evaluate the cross-sectional area (CSA) or type of muscle fibers and the diameter of myotubes. RT-qPCR, western blot, RNA sequencing (RNA-seq) and targeted metabolomic analyses were conducted to elucidate the underlying mechanisms.
Results: The results demonstrated that NTs significantly increased lean mass/body weight (p < 0.01, η2 = 0.434), the grip strength at 7 (p < 0.0001, η2 = 0.312), 9 (p < 0.001, η2 = 0.293) and 11 (p < 0.0001, η2 = 0.507) months of age and the gait speed of mice (p < 0.0001, η2 = 0.3861). The immunofluorescence staining results indicated that NTs increased the CSA of muscle fibers (p < 0.0001, η2 = 0.1081), especially for type IIb fibres. The RNA-seq results showed that NTs significantly downregulated the expression of sarcopenia-related genes (Trim63, Dkk3 and Mt1). The downregulation of Fbxo32, Trim63, Dkk3, Mt1 and p53 genes in NTs intervention group was confirmed by RT-qPCR and/or western blot (p < 0.05). Integrated analyses of RNA-seq and metabolomic showed that NTs could cause changes in metabolites, such as ketoleucine, 3-hydroxylisovalerylcarnitine and 3-methyl-2-oxovaleric acid, which could further regulate sarcopenia-related genes and inhibit protein degradation and promote protein synthesis. In vitro studies confirmed that NTs increased myotubes diameter and decreased expression of sarcopenia-related genes.
Conclusions: Our findings reveal the protective role of NTs in improving muscle protein balance during ageing, suggesting they may serve as a conditionally essential nutrient for older individuals. Further research is needed to explore the efficacy and safety of NTs supplementation for sarcopenia in humans.
Keywords: FoxO; SAMP8 mice; ageing; muscle; sarcopenia.
© 2025 The Author(s). Journal of Cachexia, Sarcopenia and Muscle published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures






References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
