

Phenotypic dichotomy in Crotalus durissus ruruima venom and potential consequences for clinical management of snakebite envenomations
- PMID: 40749023
- PMCID: PMC12327661
- DOI: 10.1371/journal.pntd.0013296
Phenotypic dichotomy in Crotalus durissus ruruima venom and potential consequences for clinical management of snakebite envenomations
Abstract
Background: Phenotypic polymorphism in rattlesnake venoms is well-documented, with a dichotomy between hemorrhagic (Type I) and neurotoxic (Type II) venoms. In South America, the Type II phenotype is predominant; however, evidence of Type I venom in Crotalus durissus ruruima raises concerns about the efficacy of the Crotalus antivenom, which is prepared only with Type II venoms. Consequently, the Bothrops-Crotalus antivenom has been proposed as an alternative treatment for envenomation by Type I venoms.
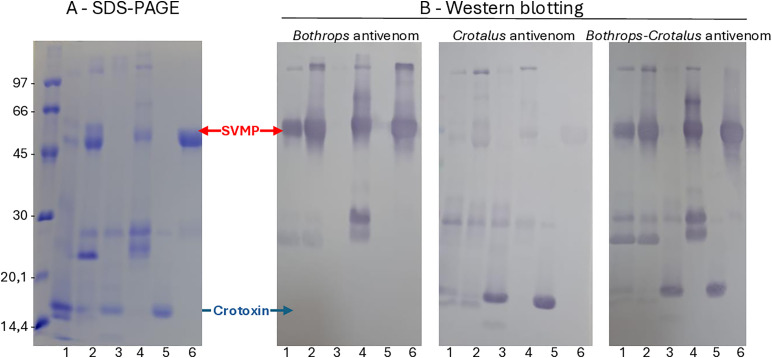
Methodology/principal findings: This study characterizes the dichotomy of C. d. ruruima venom by analyzing the structure of isoforms differentially expressed in Type I and Type II venoms, assessing their biological activities, and evaluating the implications for snakebite clinical management in Roraima State (northern Brazil). Four toxins were differentially expressed between Type I and Type II venoms: two PIII-class SVMPs, predominantly found in Type I venoms, associated with proteolytic and hemorrhagic activity; and two PLA2s, corresponding to Crotoxin A and B chains, prevalent in Type II venoms and related to elevated phospholipase A2 activity, myotoxicity, and increased lethality. The structure of Crotoxin chains was well conserved compared to C. d. terrificus Crotoxin. However, the SVMP sequences exhibited multiple substitutions in functional and immunoreactive regions compared to Bothropasin, resulting in low hemorrhagic activity and limited reactivity/neutralization by the Bothrops antivenom. Conversely, the Crotalus antivenom reacted with high antibody titer and neutralized all activities of both venom subtypes, except for the low hemorrhagic activity induced by Type I venoms.
Conclusions/significance: The efficacy of Bothrops antivenom in snakebites caused by rattlesnakes with Type I venoms remains uncertain. We advocate for a clinical study in Roraima to assess patient outcomes and benefits of Bothrops-Crotalus versus Crotalus antivenoms for these accidents. Meanwhile, administering Bothrops-Crotalus antivenom may be acceptable; however, caution is needed regarding the use of heterologous Bothrops antibodies, which have limited efficacy in treating Crotalus envenomation.
Copyright: © 2025 Colombini et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist
Figures












Similar articles
-
Ontogenetic influence in Crotalus durissus ruruima snake venom: an intermediate between north and south venom pattern of rattlesnake.Toxicon. 2025 Oct;265:108467. doi: 10.1016/j.toxicon.2025.108467. Epub 2025 Jul 3. Toxicon. 2025. PMID: 40614827
-
Comparative analysis of paralytic and myotoxic effects of Crotalus durissus terrificus venom and crotoxin from different regions of Minas Gerais, Brazil.Toxicon. 2025 Oct;265:108490. doi: 10.1016/j.toxicon.2025.108490. Epub 2025 Jul 8. Toxicon. 2025. PMID: 40639490
-
Snake venomics of the Central American rattlesnake Crotalus simus and the South American Crotalus durissus complex points to neurotoxicity as an adaptive paedomorphic trend along Crotalus dispersal in South America.J Proteome Res. 2010 Jan;9(1):528-44. doi: 10.1021/pr9008749. J Proteome Res. 2010. PMID: 19863078
-
Deciphering toxico-proteomics of Asiatic medically significant venomous snake species: A systematic review and interactive data dashboard.Toxicon. 2024 Nov 6;250:108120. doi: 10.1016/j.toxicon.2024.108120. Epub 2024 Oct 10. Toxicon. 2024. PMID: 39393539
-
Antivenom for European Vipera species envenoming.Clin Toxicol (Phila). 2017 Jul;55(6):557-568. doi: 10.1080/15563650.2017.1300261. Epub 2017 Mar 28. Clin Toxicol (Phila). 2017. PMID: 28349771
References
-
- Ministério da Saúde. Boletim epidemiológico 182023. 2023.
-
- Monteiro WM, Contreras-Bernal JC, Bisneto PF, Sachett J, Mendonça da Silva I, Lacerda M, et al. Bothrops atrox, the most important snake involved in human envenomings in the amazon: How venomics contributes to the knowledge of snake biology and clinical toxinology. Toxicon X. 2020;6:100037. doi: 10.1016/j.toxcx.2020.100037 - DOI - PMC - PubMed
-
- Sistema de Informação de Agravos de Notificação. 2017.
-
- Costa HC, Bérnils RS. Répteis do Brasil e suas unidades federativas: lista de espécies. Herpetologica Brasil. 2018;7:11–57.
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
