

Pharmacological Inhibition of c-Jun N-Terminal Kinase Activity Exacerbates Liver Damage in Schistosoma mansoni Infected Mice
- PMID: 40751475
- PMCID: PMC12317361
- DOI: 10.1111/liv.70260
Pharmacological Inhibition of c-Jun N-Terminal Kinase Activity Exacerbates Liver Damage in Schistosoma mansoni Infected Mice
Abstract
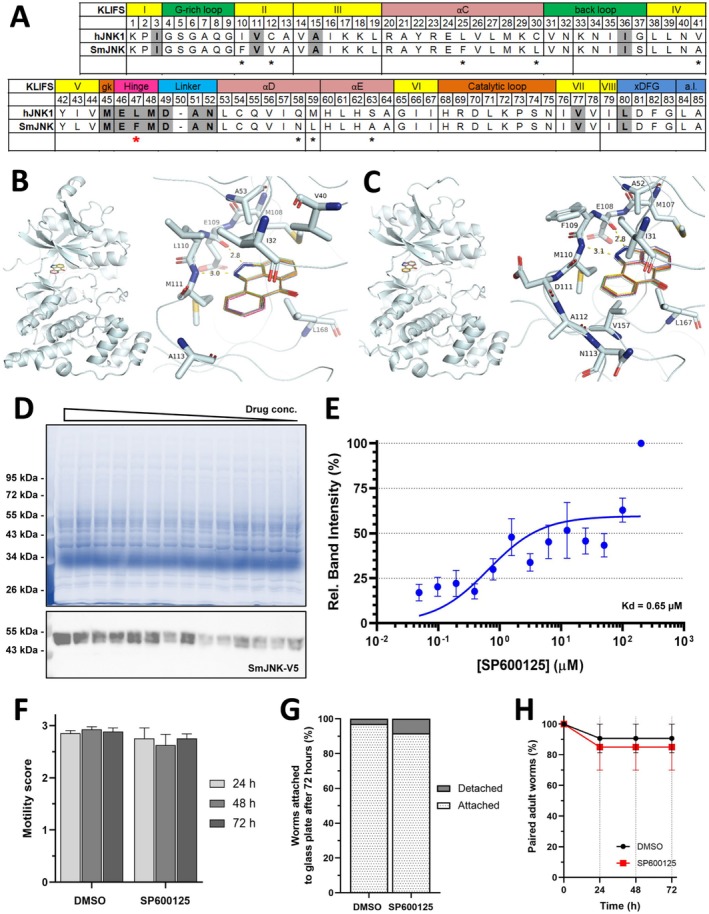
Background and aims: Schistosomiasis is a neglected tropical disease affecting more than 250 million people worldwide. Eggs of the parasitic helminth S. mansoni cause major morbidity in the liver, spleen and intestine. Of note, egg-released soluble antigens (SEA) induce the transcription factor c-Jun in hepatocytes, promoting hepatocellular cell cycle activity, proliferation and apoptosis. In this study, we analysed the hepatic effect of pharmacological inhibition of c-Jun N-terminal kinase (JNK) after infection with S. mansoni. The JNK inhibitor SP600125 was chosen because it had no effect on schistosome viability.
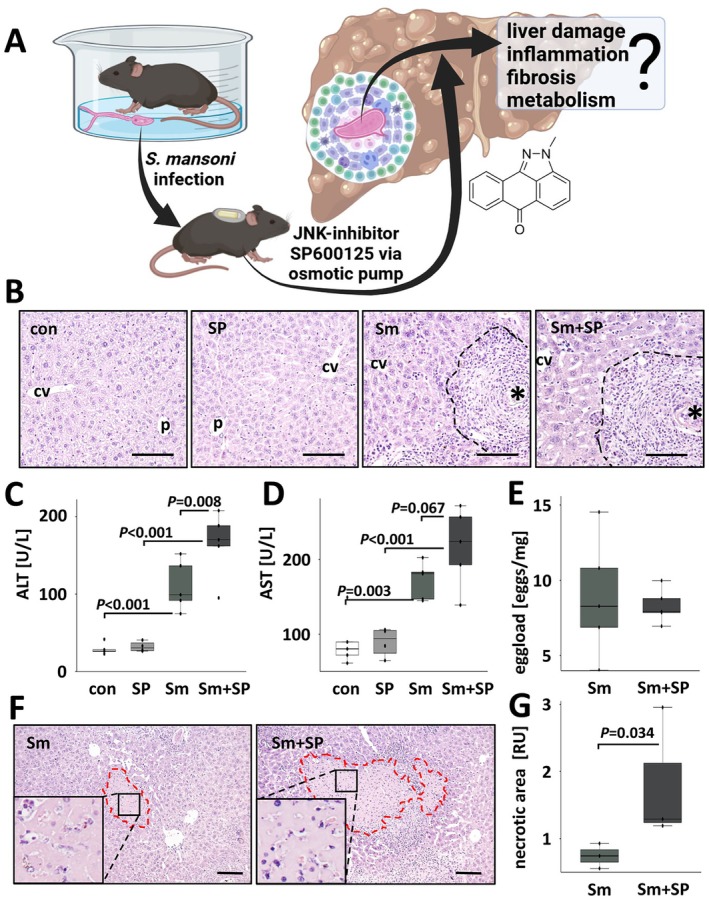
Methods: Eight-week-old male mice were infected with 100 cercariae (♂ + ♀) and 6 weeks later treated with SP600125 via a subcutaneously implanted osmotic pump over 3 weeks. Hepatic damage, inflammation, fibrosis and metabolic aspects were analysed in liver and spleen tissue as well as in serum samples.
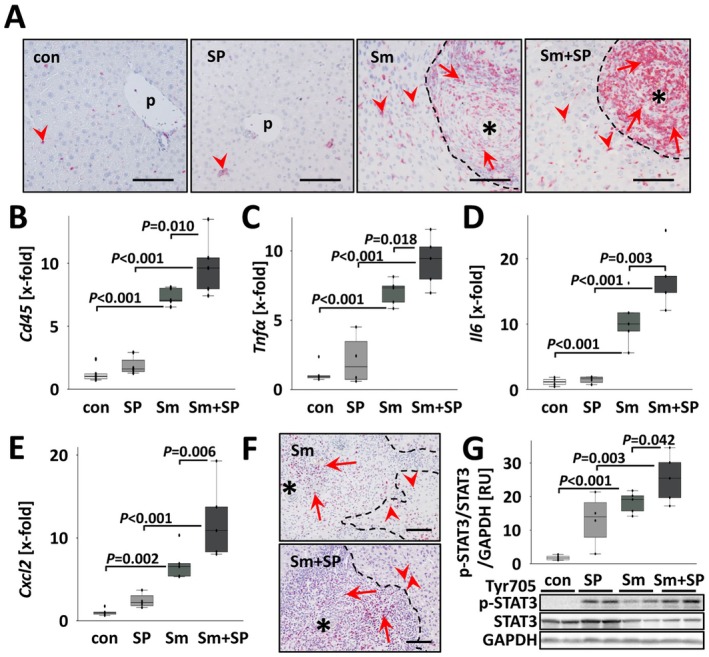
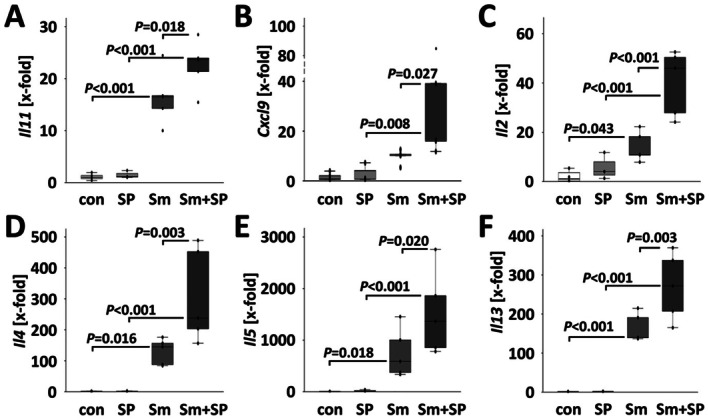
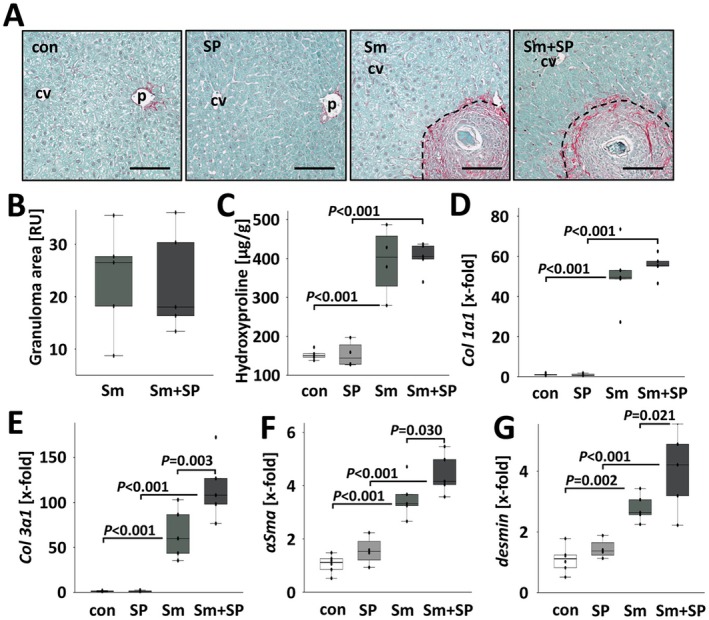
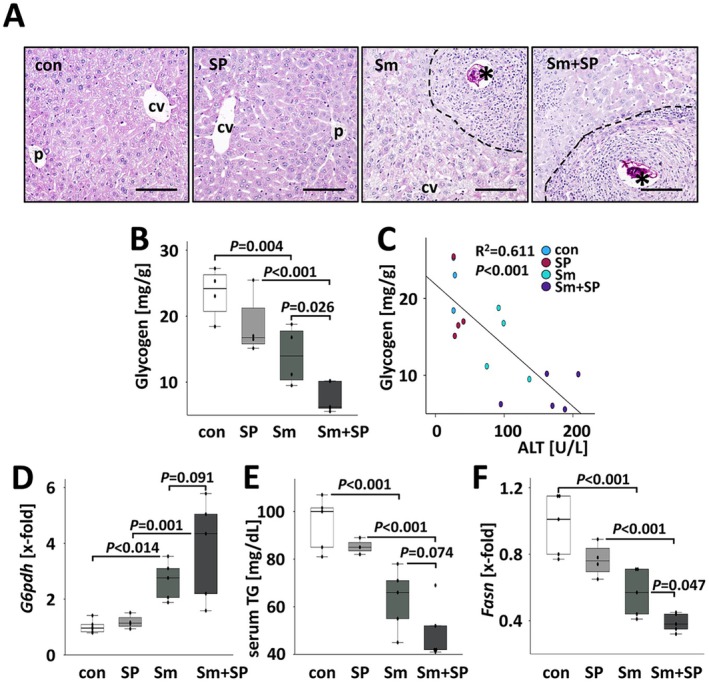
Results: JNK inhibitor-treated mice infected with S. mansoni showed a parasite-induced elevation of serum aminotransferases. Hepatic inflammation, the activation of hepatic stellate cells and metabolic exhaustion were observed in infected control mice. Additional SP600125 application almost doubled enhanced transaminases, hepatic cytokine expression, inflammation, necrosis, as well as HSC activation, and decreased glycogen stores to a minimum.
Conclusions: Our findings suggest a protective role of JNK/c-Jun-signalling in hepatic inflammation, hepatic stellate cell activation, and metabolic exhaustion during S. mansoni infection.
Keywords: SP600125; fibrosis; glycogen exhaustion; parasite; schistosomiasis.
© 2025 The Author(s). Liver International published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- WHO , “Schistosomiasis Fact Sheet, 02_2023,” 2023, https://www.who.int/news‐room/fact‐sheets/detail/schistosomiasis.
-
- McManus D. P., Dunne D. W., Sacko M., Utzinger J., Vennervald B. J., and Zhou X.‐N., “Schistosomiasis,” Nature Reviews Disease Primers 4, no. 1 (2018): 13. - PubMed
-
- Salas‐Coronas J., Bargues M. D., Lozano‐Serrano A. B., et al., “Evidence of Autochthonous Transmission of Urinary Schistosomiasis in Almeria (Southeast Spain): An Outbreak Analysis,” Travel Medicine and Infectious Disease 44 (2021): 102165. - PubMed
-
- Webster B. L., Southgate V. R., and Littlewood D. T. J., “A Revision of the Interrelationships of Schistosoma Including the Recently Described Schistosoma guineensis ,” International Journal for Parasitology 36, no. 8 (2006): 947–955. - PubMed
-
- Dunne D. W., Lucas S., Bickle Q., et al., “Identification and Partial Purification of an Antigen (Omega 1) From <styled-content style="fixed-case"> Schistosoma mansoni </styled-content> Eggs Which Is Putatively Hepatotoxic in T‐Cell Deprived Mice,” Transactions of the Royal Society of Tropical Medicine and Hygiene 75, no. 1 (1981): 54–71. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
