

Identification of the seven critical residues that control ZIKV-DENV cross-reactivity to engineer a non-cross-reactive ZIKV vaccine
- PMID: 40753572
- PMCID: PMC12440574
- DOI: 10.1016/j.celrep.2025.116098
Identification of the seven critical residues that control ZIKV-DENV cross-reactivity to engineer a non-cross-reactive ZIKV vaccine
Abstract
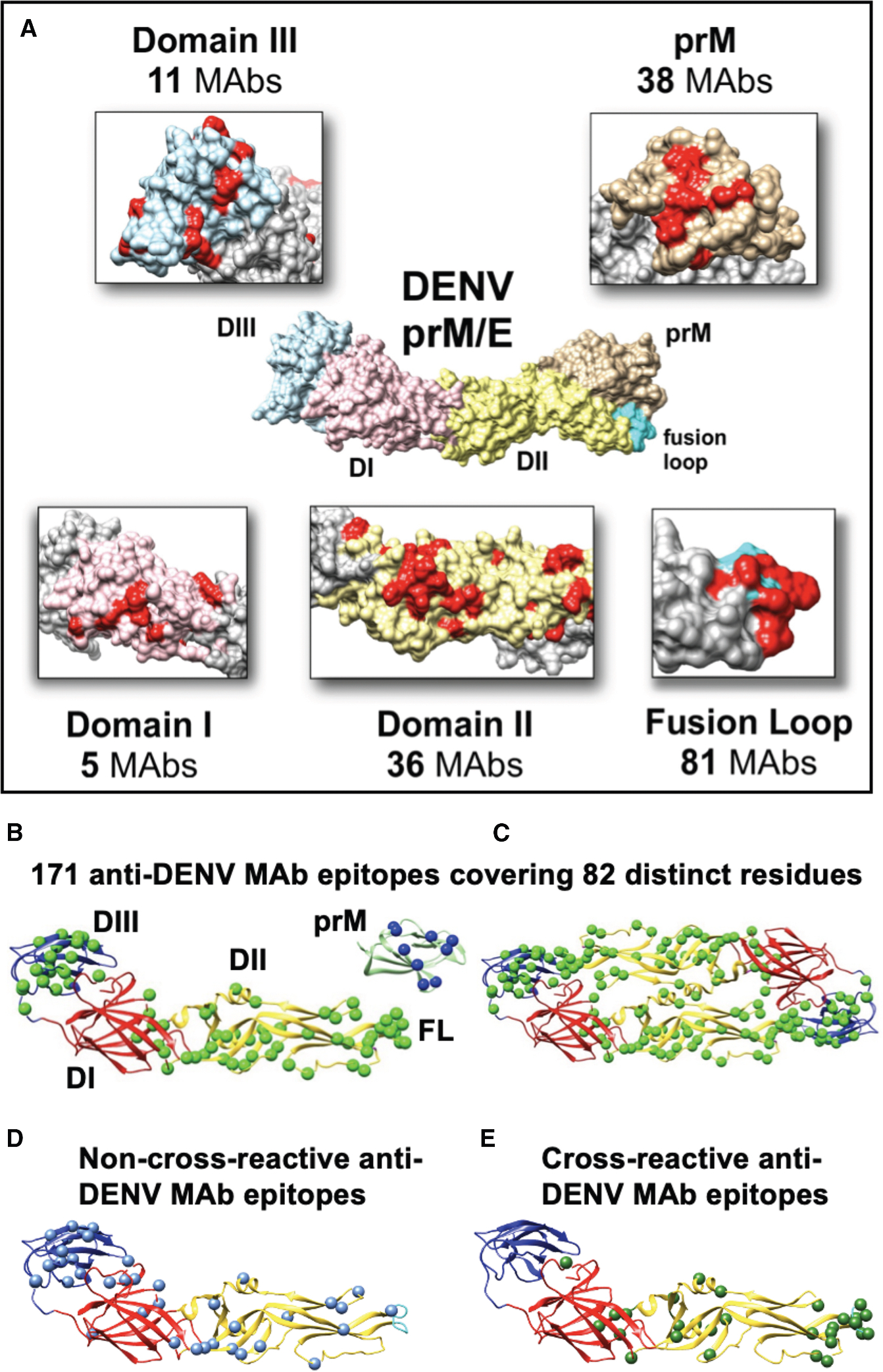
The development of a Zika virus (ZIKV) vaccine is complicated by the high homology between ZIKV and dengue virus (DENV) envelope (E) proteins, resulting in immunological cross-reactivity that can exacerbate disease through antibody-dependent enhancement (ADE). Here, we screen 121 anti-DENV monoclonal antibodies (mAbs) for cross-reactivity with ZIKV E proteins. We identify 70 cross-reactive mAbs, 66 of which have epitopes that included at least one of seven E protein residues conserved among DENV1-DENV4 and ZIKV (R73, E79, W101, L107, F108, K110, and W212), establishing these residues as the key determinants of DENV-ZIKV cross-reactivity. Using these data, we engineer a ZIKV E protein variant with 10 mutations ("ZIKVm10") that reduces cross-reactivity with DENV mAbs in vitro and minimizes the induction of anti-DENV antibodies in immunized mice. Passive serum transfer from ZIKVm10-immunized mice confers near-complete protection against lethal ZIKV challenge and reduced ADE for DENV infection, providing a pathway for improved ZIKV vaccine design.
Keywords: CP: Immunology; CP: Microbiology; antibody enhancement; antigen engineering; epitope mapping; immunogenic cross-reactivity; virus envelope.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.C.W., K.D., D.M.N., E.D., and B.J.D. are shareholders of Integral Molecular. M.J.A. is a shareholder of IBT Bioservices. M.S.D. is a consultant or member of a scientific advisory board for Inbios, Ocugen, Vir Biotechnology, Topspin Therapeutics, GlaxoSmithKline, Merck, and Moderna. The Diamond laboratory has received unrelated funding support in sponsored research agreements from Emergent BioSolutions, Moderna, Topspin Therapeutics, and Vir Biotechnology. J.E.C. has served as a consultant for Luna Labs USA, Merck Sharp & Dohme Corporation, Emergent BioSolutions, and BTG International Inc., and is a member of the scientific advisory board of Meissa Vaccines, a former member of the scientific advisory board of Gigagen (Grifols), and the founder of IDBiologics. The laboratory of J.E.C. received unrelated sponsored research agreements from AstraZeneca, Takeda Vaccines, and IDBiologics during the conduct of the study. The engineered protein described here is subject to patent 63/827,340 by A.G.-E, K.D., E.D., and B.J.D.
Figures





References
-
- Petersen E, Wilson ME, Touch S, McCloskey B, Mwaba P, Bates M, Dar O, Mattes F, Kidd M, Ippolito G, et al. (2016). Rapid Spread of Zika Virus in The Americas–Implications for Public Health Preparedness for Mass Gatherings at the 2016 Brazil Olympic Games. Int. J. Infect. Dis. 44, 11–15. 10.1016/j.ijid.2016.02.001. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
