

A CD4+ T lymphocyte-specific TCR/GSDMD/IL-2 axis facilitates antitumor immunity
- PMID: 40759573
- PMCID: PMC12321394
- DOI: 10.1172/JCI191119
A CD4+ T lymphocyte-specific TCR/GSDMD/IL-2 axis facilitates antitumor immunity
Abstract
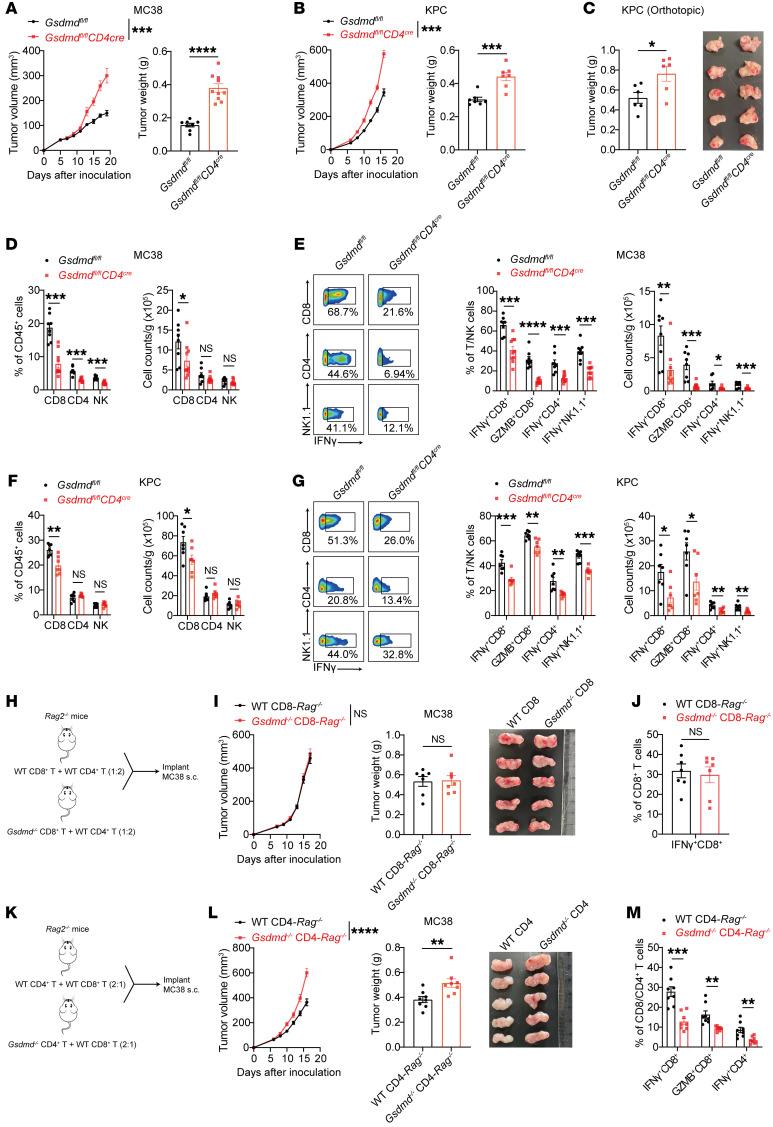
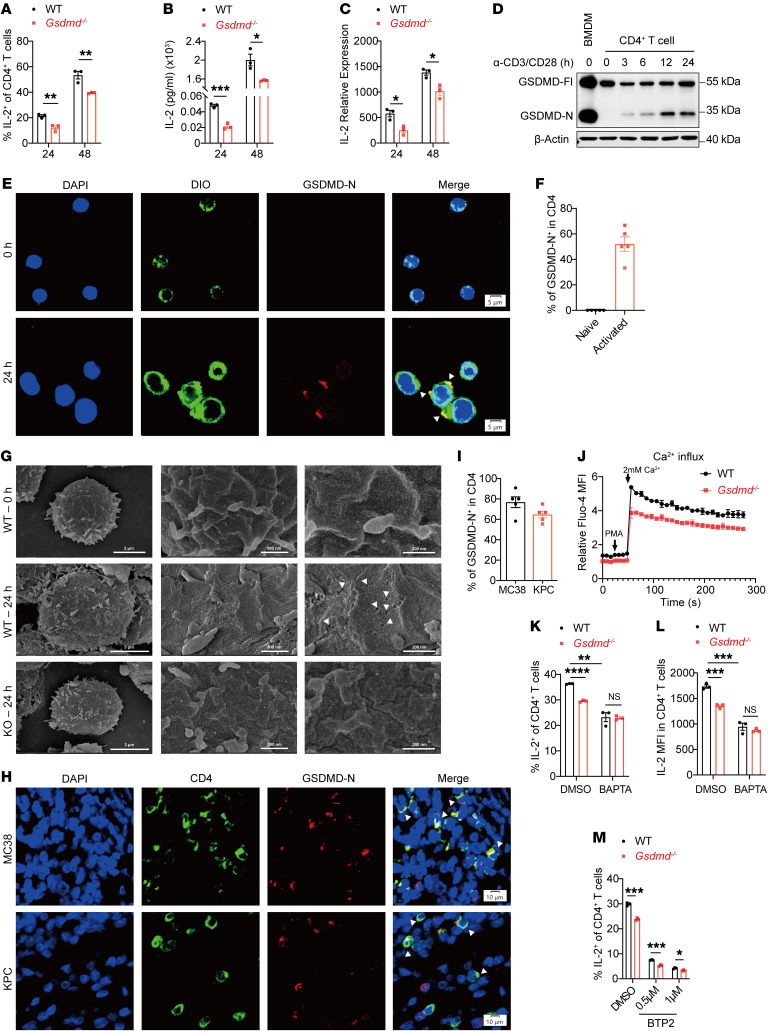
Gasdermin (GSDM) family proteins mediate tumor pyroptosis and impact cancer progression, but other than that, their involvement in the tumor immune microenvironment remains largely unknown. Here, we show that activation of GSDMD in human tumor specimens mainly occurs in tumor-infiltrating leukocytes. Significantly, GSDMD deficiency or its inactivation in CD4+ T cells disabled CD8+ T cell-mediated antitumor immunity and caused tumor outgrowth in mice. Further study uncovered that, via inducing IL-2 production, GSDMD was required for CD4+ T cells to provide help to CD8+ T cell function. Mechanistically, GSDMD was cleaved by TCR stimulation-activated caspase-8 to form GSDMD-N pores, which enhanced Ca2+ influx for IL-2 induction. Moreover, GSDMD activation and function were conserved in human CD4+ T cells and associated with favorable prognosis and improved response to anti-PD-1 immunotherapy in colonic and pancreatic cancer. We believe this study identifies a new nonpyroptotic role of GSDMD in tumor immunity, proposing GSDMD as a potential target for cancer immunotherapy.
Keywords: Calcium signaling; Cancer immunotherapy; Immunology; Oncology; T cells.
Figures





References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
