

AR to GR switch modulates differential TDO2-Kyn-AhR signalling to promote the survival and recurrence of treatment-induced dormant cells in prostate cancer
- PMID: 40759641
- PMCID: PMC12322048
- DOI: 10.1038/s41421-025-00817-w
AR to GR switch modulates differential TDO2-Kyn-AhR signalling to promote the survival and recurrence of treatment-induced dormant cells in prostate cancer
Abstract
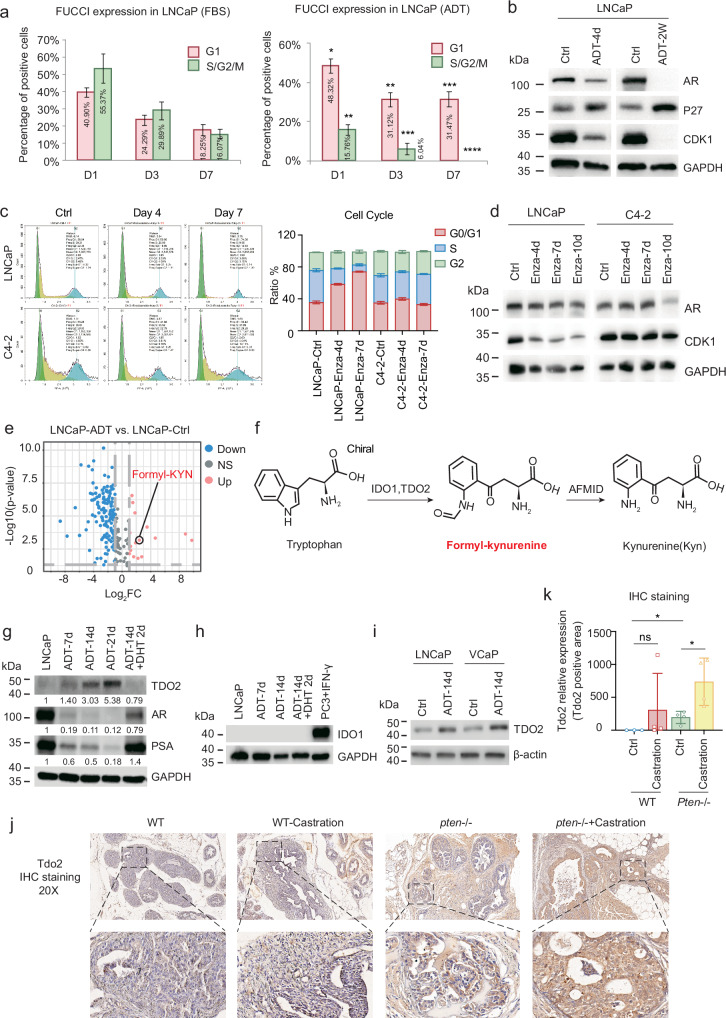
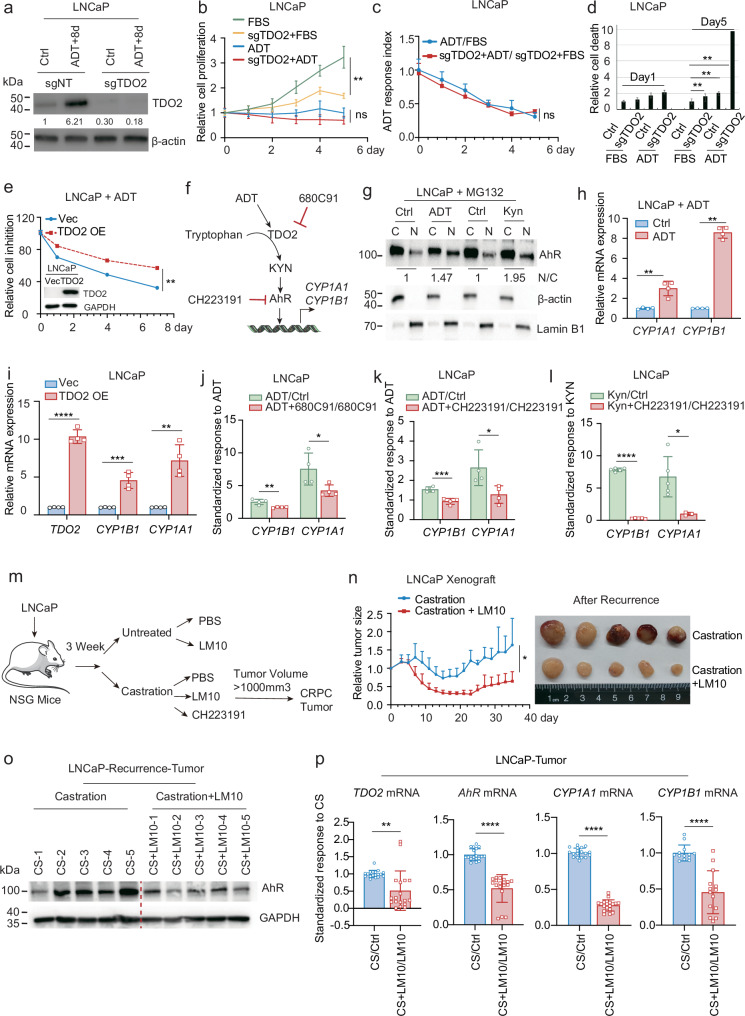
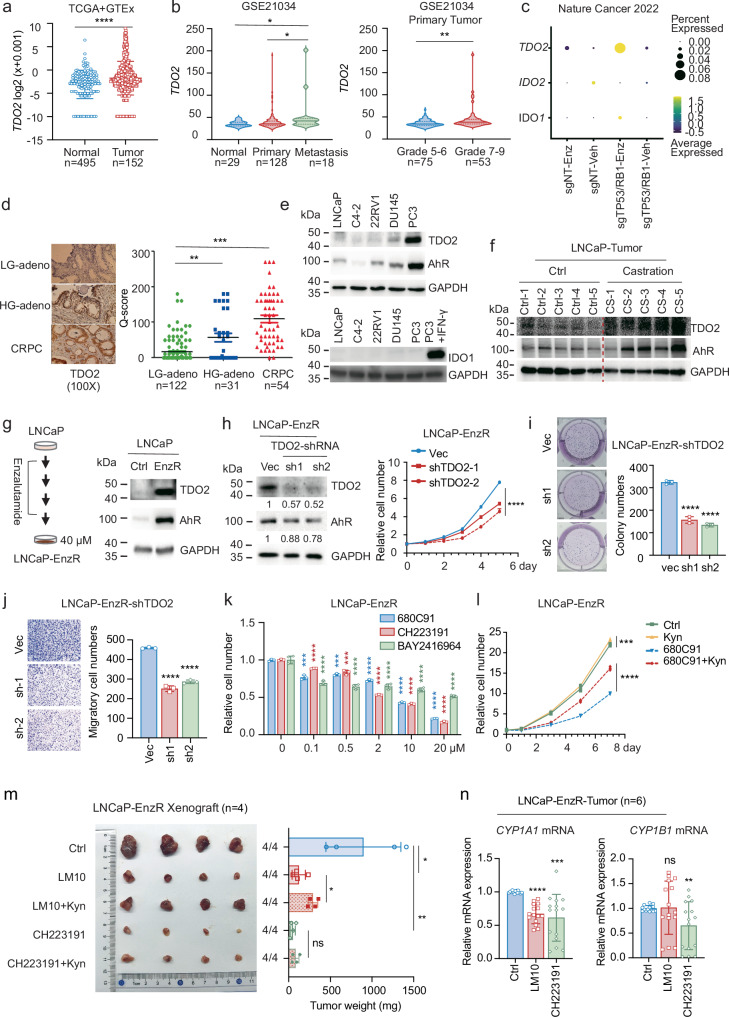
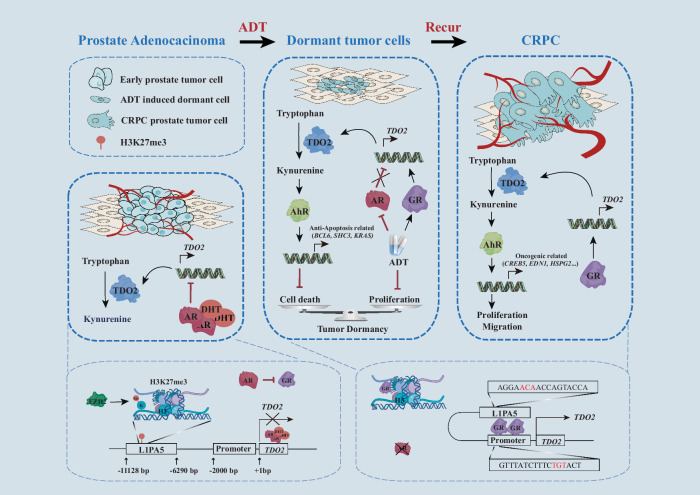
Cancer cells can be induced to dormancy initially by specific cancer therapies, but can be reactivated for subsequent relapse as therapy-resistant cancer cells. Although the treatment-induced dormancy-to-reactivation switch is an important process in tumour spread and recurrence, little is known about the underlying molecular mechanisms, particularly the metabolic underpinnings. In this study, we demonstrated that the tryptophan catabolism-related tryptophan 2,3-dioxygenase (TDO2) -kynurenine (Kyn) -aryl hydrocarbon receptor (AhR) signalling axis was responsible for both sustaining the survival of dormant prostate cancer cells induced by androgen deprivation therapy (ADT) and promoting the reactivation of dormant cells and their recurrent outgrowth, which facilitated the development of therapeutic resistance by allowing the dormancy-to-reactivation switch. Mechanistically, we found that ADT upregulated the expression of TDO2 to produce Kyn, which activated AhR and maintained the survival of ADT-induced dormant cells. Interestingly, the switch of transcription factors from the androgen receptor (AR) to the glucocorticoid receptor (GR) modulated the persistent expression of TDO2 and promoted the reactivation of dormant cells through the same TDO2-Kyn-AhR signalling axis. Additionally, tumour recurrence following ADT was delayed by pharmacological suppression of TDO2-Kyn-AhR signalling with a TDO2 inhibitor or an AhR inhibitor. In summary, we describe a signalling circuit mediated by tryptophan metabolism for regulating tumour cell dormancy and recurrence and propose TDO2 as a new target for the treatment of androgen-sensitive prostate cancer patients in combination with ADT.
© 2025. The Author(s).
Conflict of interest statement
Conflict of interest: The authors declare no competing interests. Ethics: This study was carried out in accordance with the recommendations of the Southern University of Science and Technology Laboratory Animal Welfare Ethics Review Committee.
Figures


Similar articles
-
Dual inhibition of IDO1/TDO2 enhances anti-tumor immunity in platinum-resistant non-small cell lung cancer.Cancer Metab. 2023 May 24;11(1):7. doi: 10.1186/s40170-023-00307-1. Cancer Metab. 2023. PMID: 37226257 Free PMC article.
-
IDO2-AhR axis as central regulator of the kynurenine pathway in glioblastoma.J Neurooncol. 2025 Sep;174(3):721-731. doi: 10.1007/s11060-025-05106-w. Epub 2025 Jun 5. J Neurooncol. 2025. PMID: 40471422 Free PMC article.
-
TDO2 Deficiency Exacerbates the Immune Rejection Response in Rat Liver Transplantation via the Kyn-AHR Axis.Transplantation. 2025 Aug 1;109(8):e386-e399. doi: 10.1097/TP.0000000000005386. Epub 2025 Apr 1. Transplantation. 2025. PMID: 40164990
-
EAU guidelines on prostate cancer. Part II: Treatment of advanced, relapsing, and castration-resistant prostate cancer.Eur Urol. 2014 Feb;65(2):467-79. doi: 10.1016/j.eururo.2013.11.002. Epub 2013 Nov 12. Eur Urol. 2014. PMID: 24321502
-
Systemic treatments for metastatic cutaneous melanoma.Cochrane Database Syst Rev. 2018 Feb 6;2(2):CD011123. doi: 10.1002/14651858.CD011123.pub2. Cochrane Database Syst Rev. 2018. PMID: 29405038 Free PMC article.
References
-
- Phan, T. G. & Croucher, P. I. The dormant cancer cell life cycle. Nat. Rev. Cancer20, 398–411 (2020). - PubMed
Grants and funding
- #82173131/National Natural Science Foundation of China (National Science Foundation of China)
- #JCYJ20240813095300002/Shenzhen Science and Technology Innovation Commission
- #2023A1515010124/Natural Science Foundation of Guangdong Province (Guangdong Natural Science Foundation)
- #2024A1515012783/Natural Science Foundation of Guangdong Province (Guangdong Natural Science Foundation)
LinkOut - more resources
Full Text Sources
Research Materials
