

AgRP neuron activity enhances reward-related consummatory behaviors during energy deficit in mice
- PMID: 40760098
- PMCID: PMC12321993
- DOI: 10.1038/s42003-025-08620-9
AgRP neuron activity enhances reward-related consummatory behaviors during energy deficit in mice
Abstract
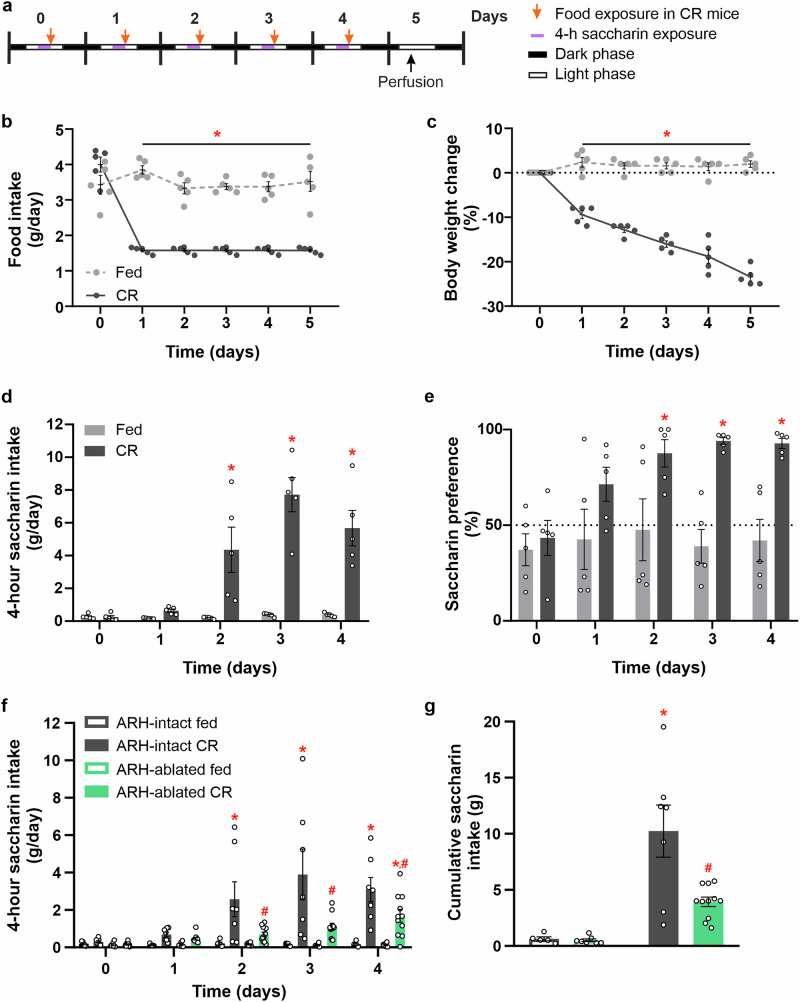
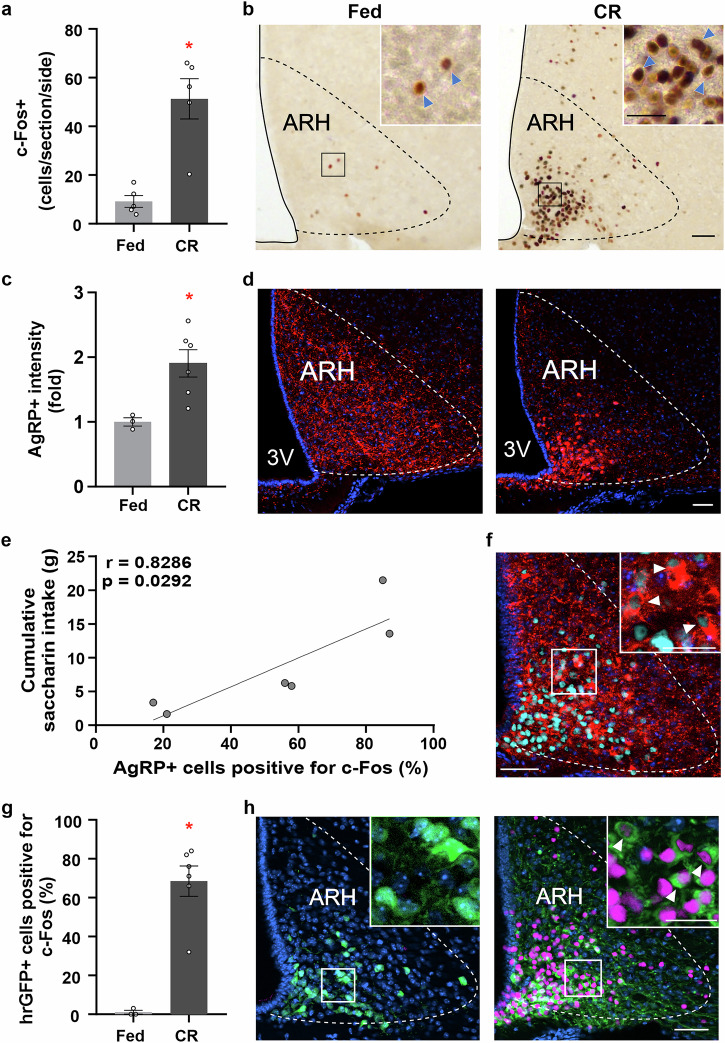
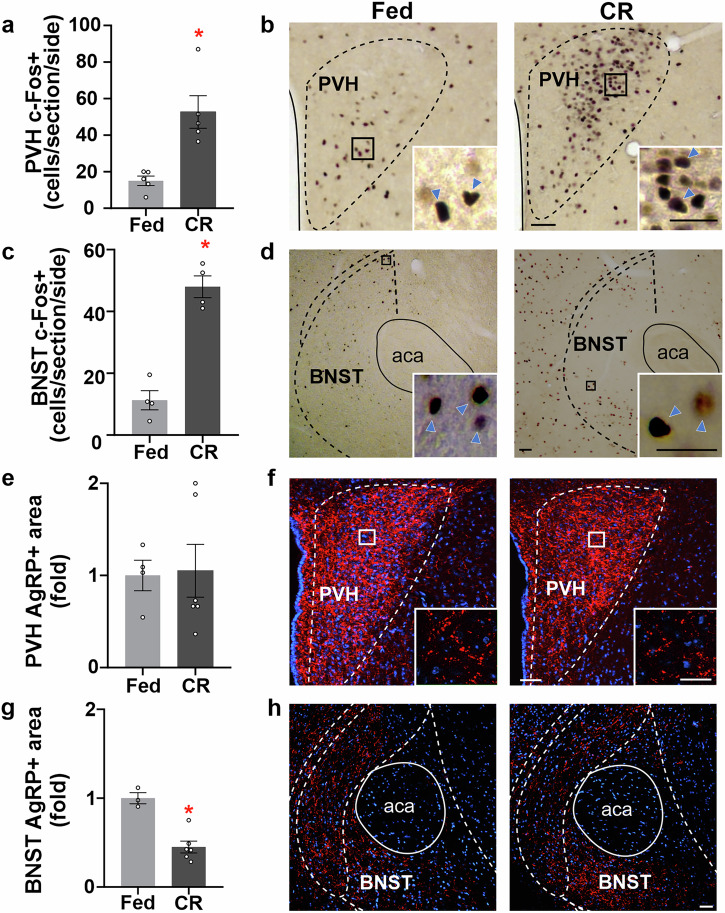
Hunger enhances the consumption of rewarding foods, but the neurobiological basis of this adaptation remains unclear. We hypothesize that agouti-related protein (AgRP) neurons in the hypothalamic arcuate nucleus (ARH) promote the consumption of rewarding stimuli under calorie restriction, independent of caloric content. To test this, we study mice fed 40% of their average daily intake and exposed daily to the non-caloric sweetener saccharin before feeding. We show that calorie-restricted (CR) mice increase saccharin intake before each restricted feeding event and that this response requires ARH integrity. CR mice exhibit activation of AgRP neurons and their brain targets without significant changes in AgRP fiber density. Furthermore, satiated mice increase saccharin intake following chemogenetic activation of AgRP neurons, whereas CR mice with selective chemogenetic inhibition of AgRP neurons show reduced saccharin intake. Thus, we conclude that AgRP neuron activation enhances the consumption of a purely rewarding stimulus in CR mice. These findings contribute to our understanding of how the brain shapes food choices under conditions of energy deficit and could be important for managing food consumption during dieting or in eating disorders.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics approval: All experiments received approval from the Institutional Animal Care and Use Committees of the IMBICE (21-0430 A) and the Federal University of São Paulo (12/2017). Consent to publish and consent to participate: The study does not involve human subjects and the manuscript contains any individual person’s data in any form.
Figures

References
-
- Luquet, S., Perez, F. A., Hnasko, T. S. & Palmiter, R. D. NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science310, 683–685 (2005). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
