

MR1-ligand cross-linking identifies vitamin B6 metabolites as TCR-reactive antigens
- PMID: 40763744
- PMCID: PMC12461623
- DOI: 10.1016/j.crmeth.2025.101120
MR1-ligand cross-linking identifies vitamin B6 metabolites as TCR-reactive antigens
Abstract
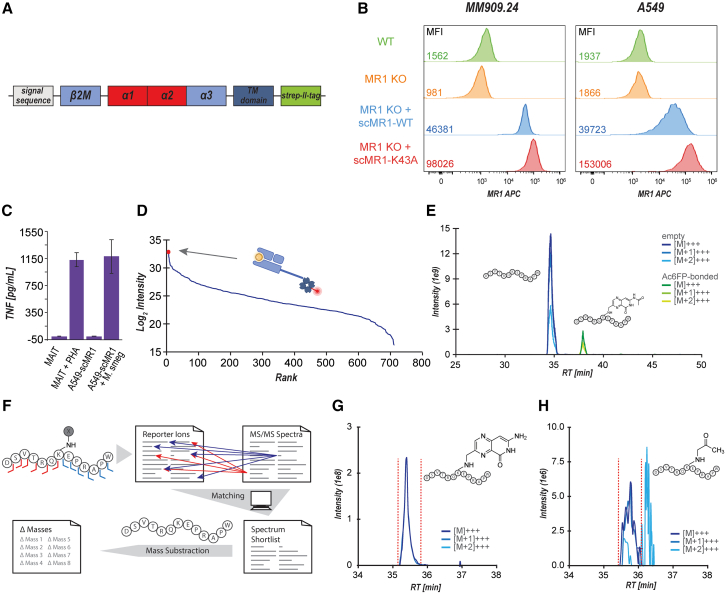
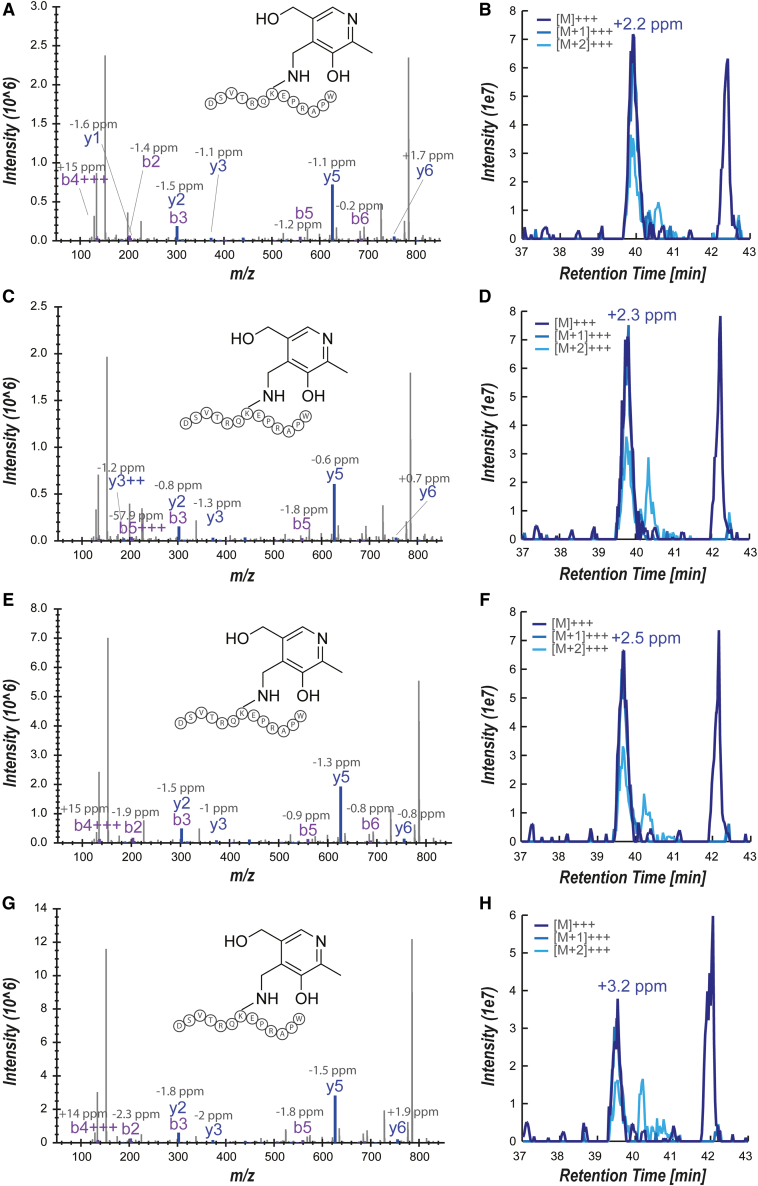
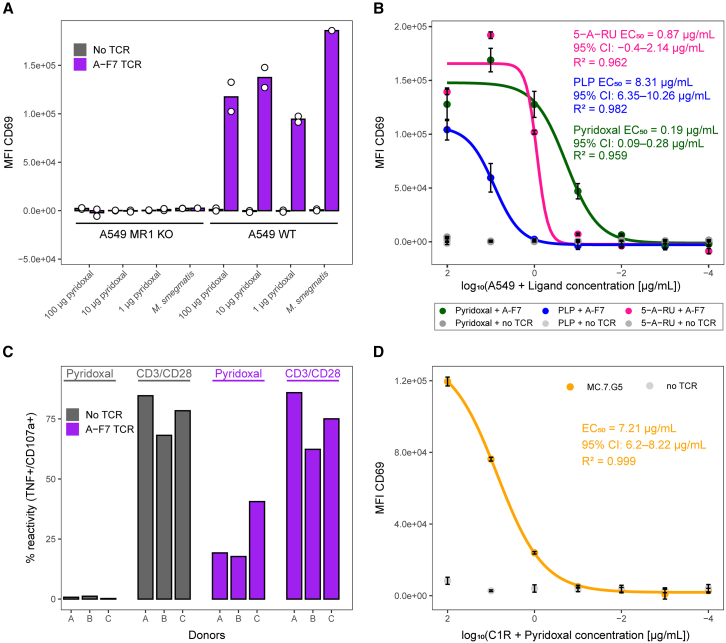
Major histocompatibility complex class I-related protein 1 (MR1) plays a central role in the immune recognition of infected cells and can mediate T cell detection of cancer. Knowledge of the nature of the ligands presented by MR1 is still sparse and has been limited by a lack of efficient approaches for MR1 ligand discovery. Here, we present a cross-linking strategy to investigate Schiff base-bound MR1 ligands. Our methodology employs reductive amination to stabilize the labile Schiff base bond between MR1 and its ligand, allowing for the detection of ligands as covalent MR1 adducts by mass spectrometry-based proteomics. We apply our approach to identifying vitamin B6 vitamers pyridoxal and pyridoxal 5'-phosphate (PLP) as MR1 ligands and show that both compounds are recognized by T cells expressing either A-F7, a mucosal-associated invariant T (MAIT) cell T cell receptor (TCR), or MC.7.G5, an MR1-restricted TCR reported to recognize cancer cells, highlighting them as immunogenic MR1 ligands.
Keywords: CP: Immunology; MAIT; MR1; T cell; TCR; antigen presentation; cross-linking; mass spectrometry; metabolite antigens; pyridoxal; vitamin B6.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests F.M., M.v.E., R.M.G., H.S., J.D., J.D.S., and N.T. were employees of Enara Bio at the time of this study and were in receipt of salary and stock options in Enara Bio Ltd. For parts of the study, T.S., E.B., and S.H. were academic postdoctoral researchers fully or partly funded by Enara Bio Ltd. A.K.S. and G.D. have patents granted and pending on T cell recognition.
Figures




References
-
- Crowther M.D., Dolton G., Legut M., Caillaud M.E., Lloyd A., Attaf M., Galloway S.A.E., Rius C., Farrell C.P., Szomolay B., et al. Genome-wide CRISPR-Cas9 screening reveals ubiquitous T cell cancer targeting via the monomorphic MHC class I-related protein MR1. Nat. Immunol. 2020;21:178–185. doi: 10.1038/s41590-019-0578-8. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
