

P-Rex2 suppresses glucose uptake into liver and skeletal muscle through different adaptor functions
- PMID: 40764335
- PMCID: PMC12325715
- DOI: 10.1038/s41598-025-01720-w
P-Rex2 suppresses glucose uptake into liver and skeletal muscle through different adaptor functions
Abstract
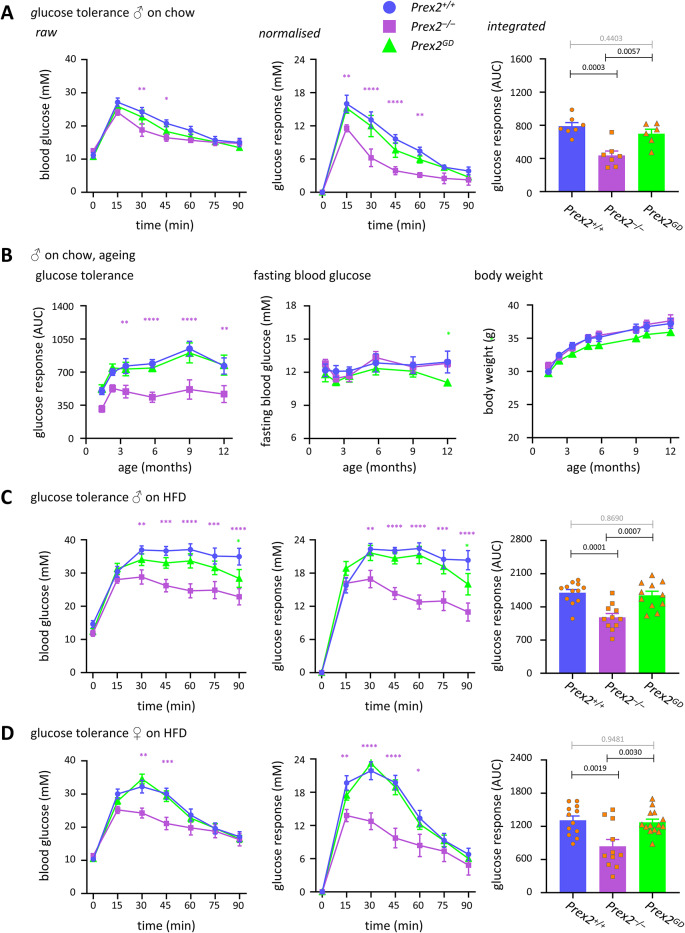
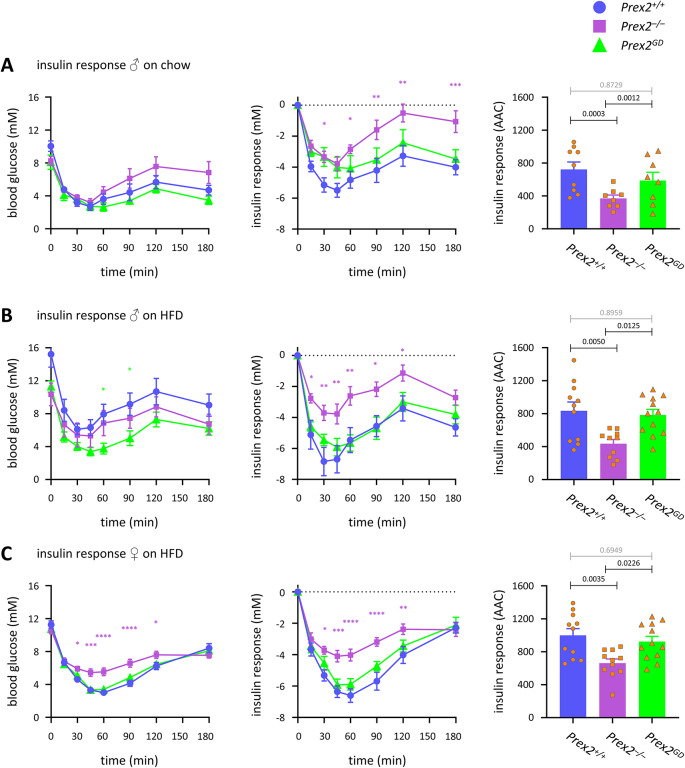
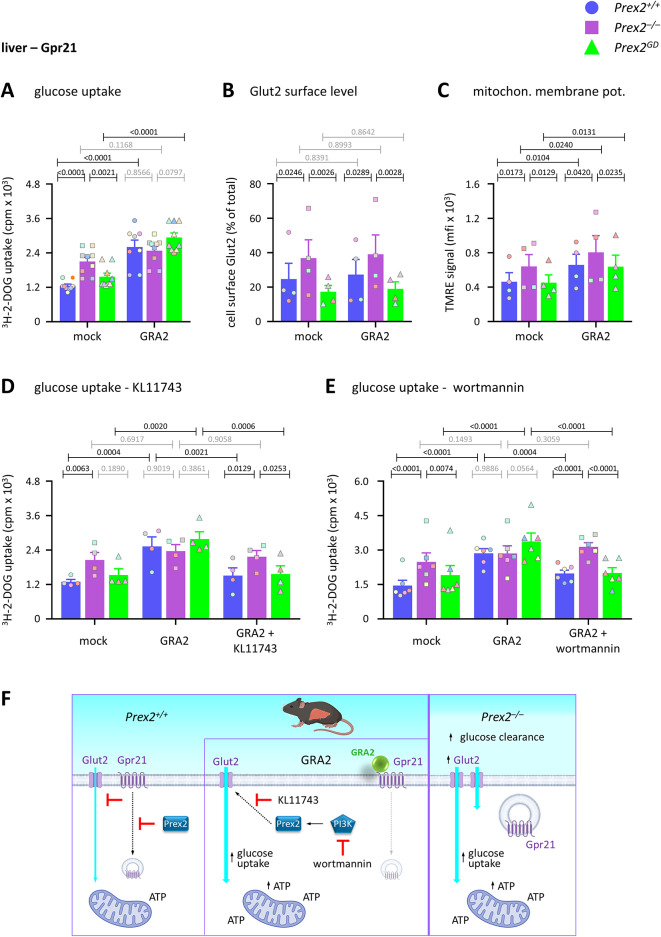
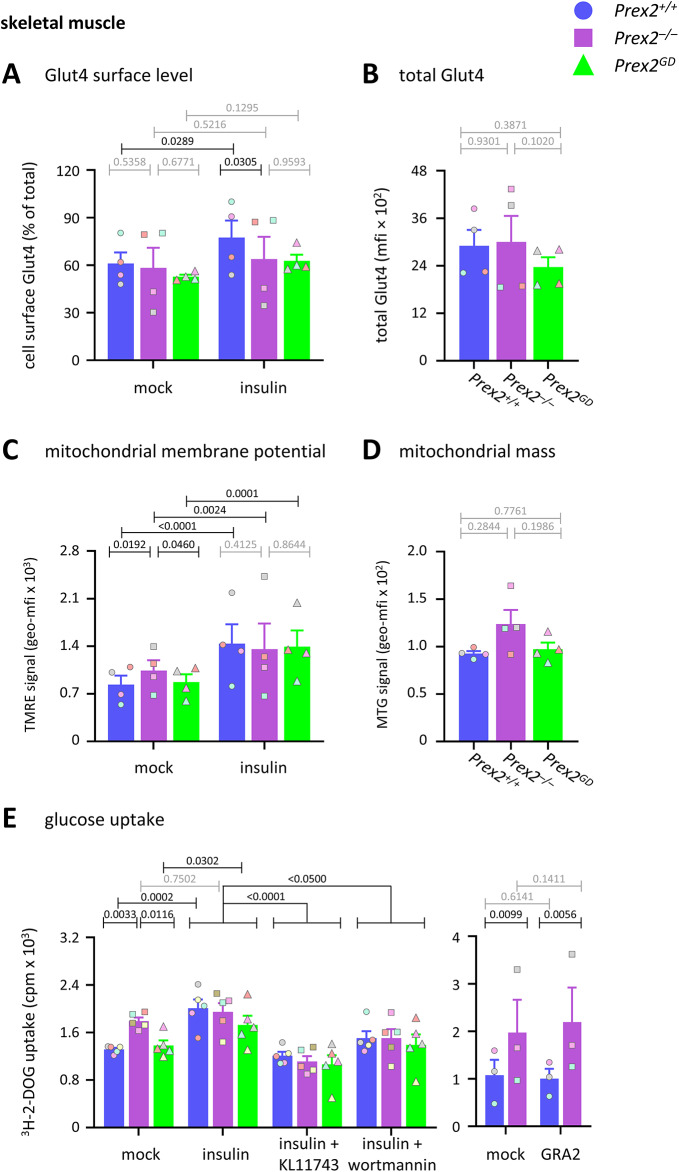
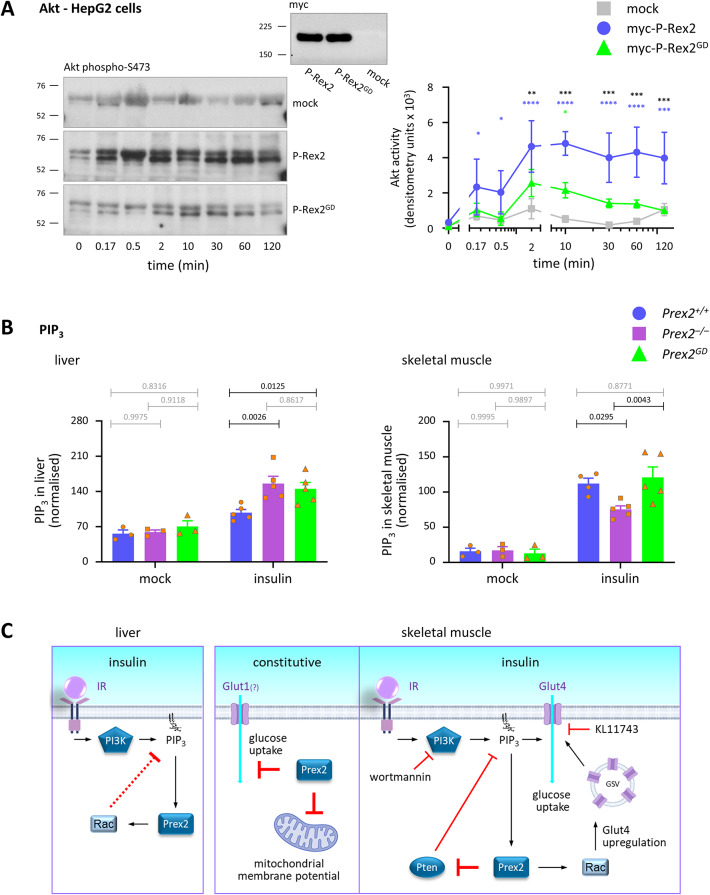
P-Rex2 is a Rac guanine-nucleotide factor (Rac-GEF) that controls glucose homeostasis. This role is thought to be mediated through its adaptor function inhibiting Pten rather than through its Rac-GEF activity, but this remains to be demonstrated. To examine this question, we have investigated the roles of P-Rex2 in glucose homeostasis using Prex2-/- and catalytically-inactive Prex2GD mice. We show that P-Rex2 is required for insulin sensitivity but limits glucose clearance, suppressing glucose uptake into liver and skeletal muscle independently of its catalytic activity. In hepatocytes, P-Rex2 suppresses Glut2 cell surface levels, mitochondrial membrane potential and mitochondrial ATP production. We identify the orphan GPCR Gpr21 as a P-Rex2 target and propose that P-Rex2 limits hepatic glucose clearance by controlling Gpr21 trafficking. In skeletal muscle cells, P-Rex2 suppresses glucose uptake through a separate adaptor function, independently of Gpr21. Additionally, P-Rex2 suppresses insulin secretion by pancreatic islets and plasma insulin levels. Finally, P-Rex2 plays distinct Rac-GEF activity dependent and independent roles in PIP3 production in liver and skeletal muscle, respectively. Together, our study identifies complex roles of P-Rex2 in glucose homeostasis, mediated through largely GEF-activity independent mechanisms which include the GPCR Gpr21 in hepatocytes and but are not obviously linked to the regulation of Pten.
Keywords: G protein-coupled receptor (GPCR); Glucose homeostasis; Gpr21; Guanine-nucleotide exchange factor (GEF); Liver; Mitochondria; P-Rex1; P-Rex2; Skeletal muscle.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures


References
-
- Gerich, J. E. Physiology of glucose homeostasis. Diabetes Obes Metab2, 345–350. 10.1046/j.1463-1326.2000.00085.x (2000). - PubMed
-
- Ueda, S., Kataoka, T. & Satoh, T. Activation of the small GTPase Rac1 by a specific guanine-nucleotide-exchange factor suffices to induce glucose uptake into skeletal-muscle cells. Biol Cell100, 645–657. 10.1042/BC20070160 (2008). - PubMed
MeSH terms
Substances
Grants and funding
- BB/P013384/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P013384/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P013384/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P013384/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P013384/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
