

This is a preprint.
Structures of dynamic interactors at native proteasomes by PhIX-MS and cryoelectron microscopy
- PMID: 40766414
- PMCID: PMC12324443
- DOI: 10.1101/2025.07.31.667872
Structures of dynamic interactors at native proteasomes by PhIX-MS and cryoelectron microscopy
Abstract
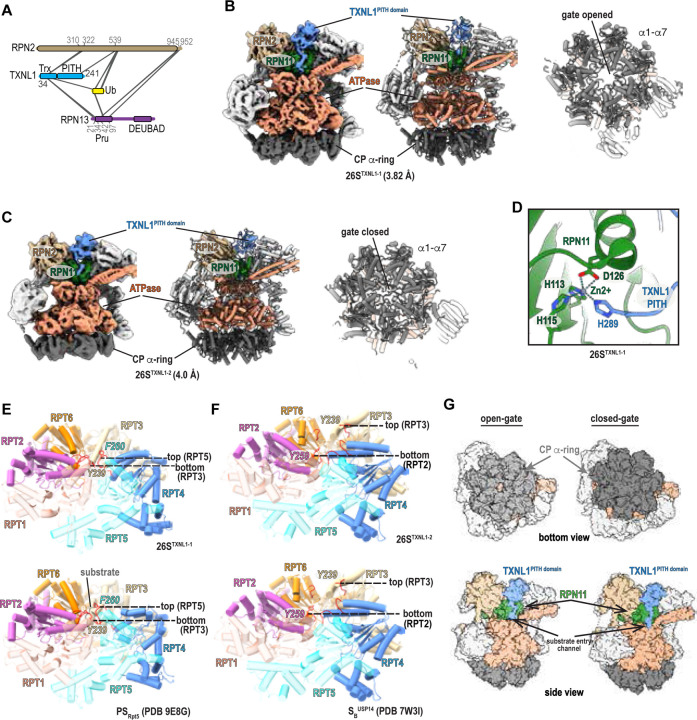
Proteasome function depends on a network of transient interactions that remain structurally and functionally unresolved. We developed PhIX-MS (Photo-induced In situ Crosslinking-Mass Spectrometry), a structural proteomics workflow that stabilizes transient interactions in cells by UV-activated crosslinking to capture topological information. Applying PhIX-MS with cryo-electron microscopy (cryo-EM), we mapped redox sensor TXNL1 at the proteasome regulatory particle (RP), placing its PITH domain above deubiquitinase RPN11 and resolving its dynamic thioredoxin domain near RPN2/PSMD1 and RPN13/ADRM1, ideally located to reduce substrates prior to proteolysis. We also resolved chaperone PSMD5 bound to RP without the proteolytic core particle (CP) where its C-terminus inserts into the ATPase pore blocking CP binding. PhIX-MS and AlphaFold modeling tether ubiquitin ligase UBE3C/Hul5 along the RP placing its catalytic site above the RPN11 active site, enabling their coupled activities. Our integrative approach enables the localization of native, low-affinity protein interactions and is broadly applicable to dynamic macromolecular assemblies.
Keywords: PSMD5; TXNL1; UBE3C; in situ mass spectrometry; proteasome.
Conflict of interest statement
Declarations of Interest The authors declare no competing interests.
Figures





References
-
- Buel G.R., Lu X., and Walters K.J. (2022). Exploiting the Proteasome for Disease Treatment. In Protein Homeostasis in Drug Discovery, pp. 135–177. 10.1002/9781119774198.ch4. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous