

Genome-wide CRISPR screen reveals key role of sialic acids in PEDV and porcine coronavirus infections
- PMID: 40767522
- PMCID: PMC12421841
- DOI: 10.1128/mbio.01628-25
Genome-wide CRISPR screen reveals key role of sialic acids in PEDV and porcine coronavirus infections
Abstract
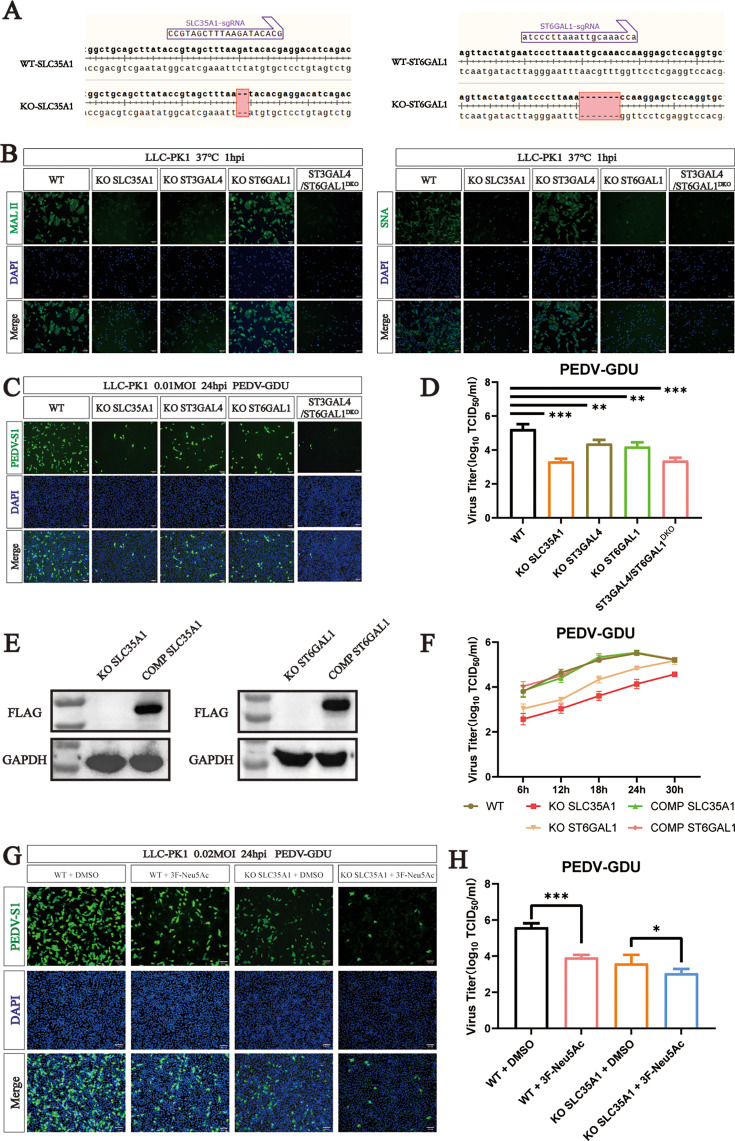
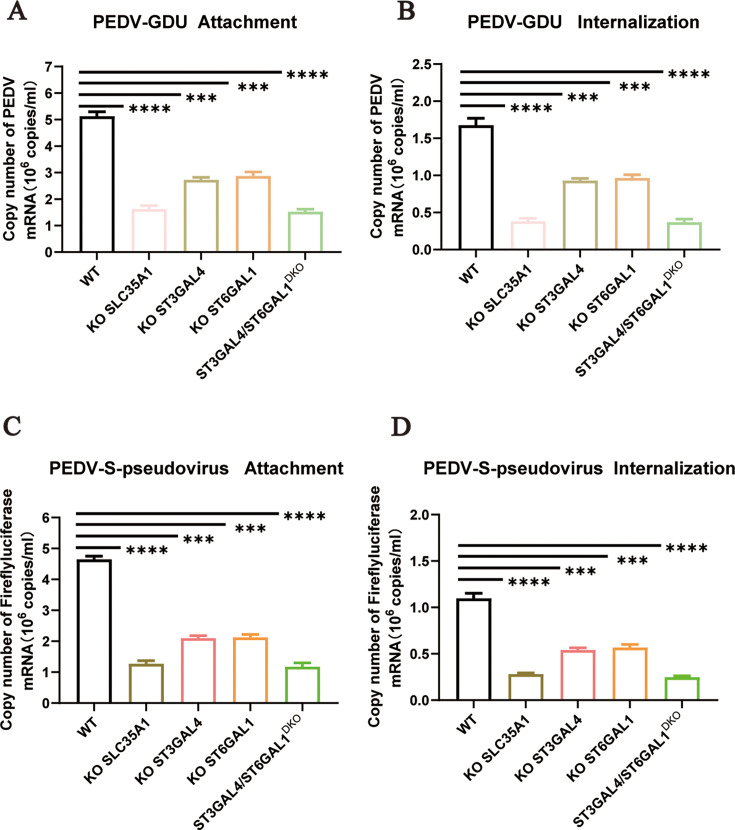
Porcine epidemic diarrhea virus (PEDV) is a globally distributed alphacoronavirus with economic importance that can cause severe watery diarrhea and even death in piglets. To identify host factors essential for PEDV infection, we performed a genome-wide CRISPR/Cas9 screen in human hepatocellular carcinoma cells (Huh7) using the highly virulent PEDV GIIb strain GDU. Several genes involved in the sialic acid and heparan sulfate biosynthesis pathway and cholesterol metabolism were highly enriched following PEDV selection. We validated that the host factor ST3 beta-galactoside alpha-2,3-sialyltransferase 4 (ST3GAL4), which catalyzes the transfer of sialic acid to sugar chains via α2,3-linked linkages, is important for PEDV infection. To systematically investigate the role of sialic acid in PEDV infection, we knocked out genes related to sialic acid synthesis. This led to a reduced abundance of sialic acid on the cell surface, which in turn inhibited PEDV adsorption and internalization. Furthermore, we found that both α2,3-linked and α2,6-linked sialic acids can serve as cellular attachment factors for PEDV. We conducted a glycan microarray screen to determine which sialoglycans are preferred by the PEDV spike protein. The results revealed that PEDV favors binding to α2,3-sialoglycans. Additionally, we found that not only current circulating PEDV strains but also other porcine coronaviruses rely on sialic acid for efficient infection. Collectively, our findings provide insights into critical host factors involved in PEDV infection and demonstrate that disrupting genes involved in sialic acid biosynthesis negatively affects the infectivity of multiple porcine enteric coronaviruses.IMPORTANCEA wide range of viruses utilize sialic acid as receptors. Sialic acid binding may serve as a key determinant of viral host range. Different viruses exhibit distinct preferences for specific types of sialic acid linkages. However, it remains unclear which specific subtypes of sialic acid are utilized during PEDV infection. In this study, we performed CRISPR-based genome-wide knockout screening and identified ST3GAL4 as a key host factor for PEDV infection. Furthermore, we found that both α2,3-linked and α2,6-linked sialic acids can function as attachment factors for PEDV infection. A glycan microarray screen revealed that PEDV S1 shows the strongest binding preference for α2,3-linked and α2,8-linked sialosides. Sialic acids were also implicated in infections by other porcine enteric coronaviruses. Overall, our findings advance our understanding of viral entry mechanisms of PEDV and other swine coronaviruses and may provide avenues for designing antiviral strategies.
Keywords: PEDV; SLC35A1; ST3GAL4; ST6GAL1; coronaviruses; sialic acid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Chen Q, Li G, Stasko J, Thomas JT, Stensland WR, Pillatzki AE, Gauger PC, Schwartz KJ, Madson D, Yoon K-J, Stevenson GW, Burrough ER, Harmon KM, Main RG, Zhang J. 2014. Isolation and characterization of porcine epidemic diarrhea viruses associated with the 2013 disease outbreak among swine in the United States. J Clin Microbiol 52:234–243. doi: 10.1128/JCM.02820-13 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
