

Immiscible proteins compete for RNA binding to order condensate layers
- PMID: 40768359
- PMCID: PMC12338069
- DOI: 10.1073/pnas.2504778122
Immiscible proteins compete for RNA binding to order condensate layers
Abstract
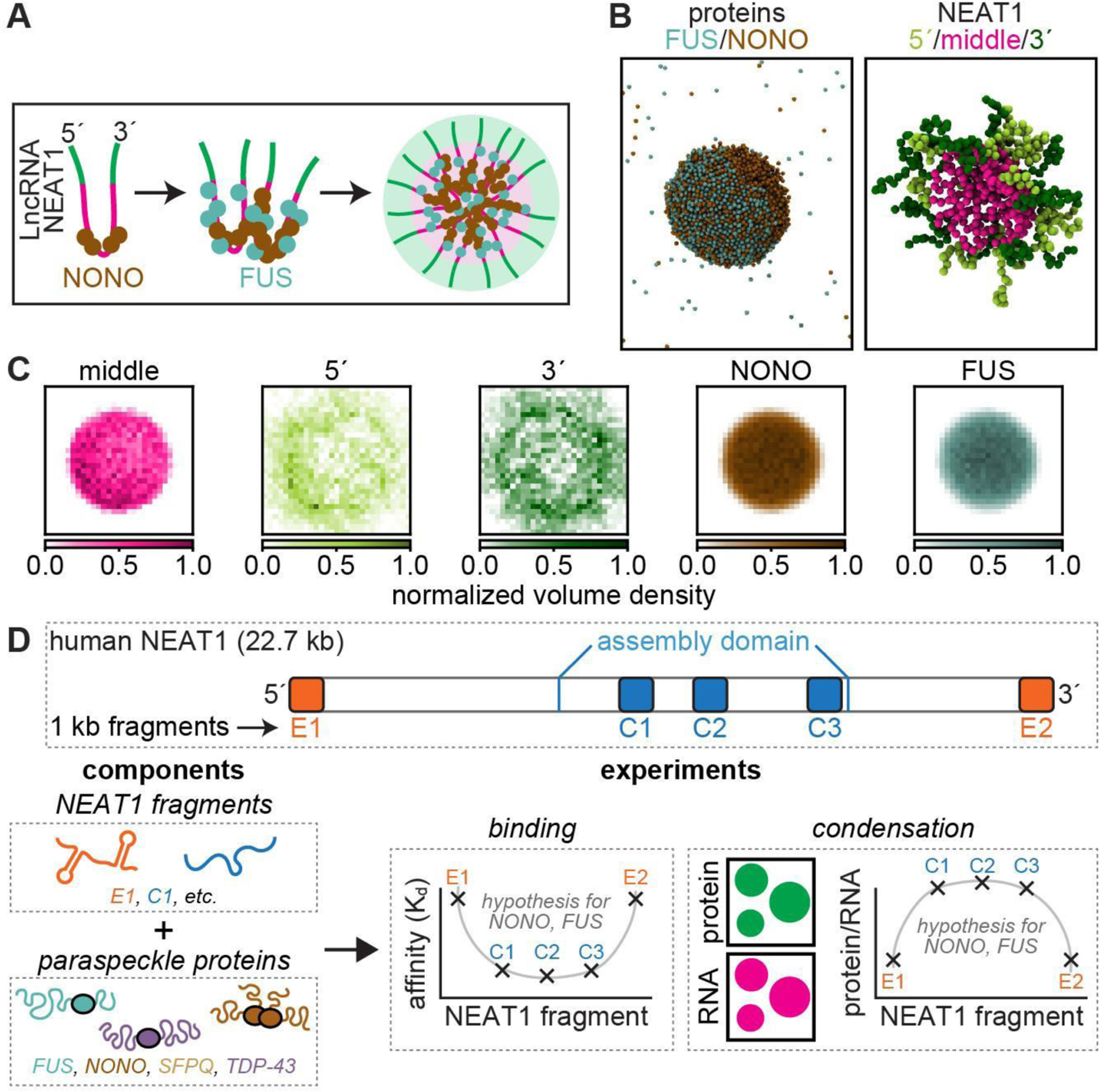
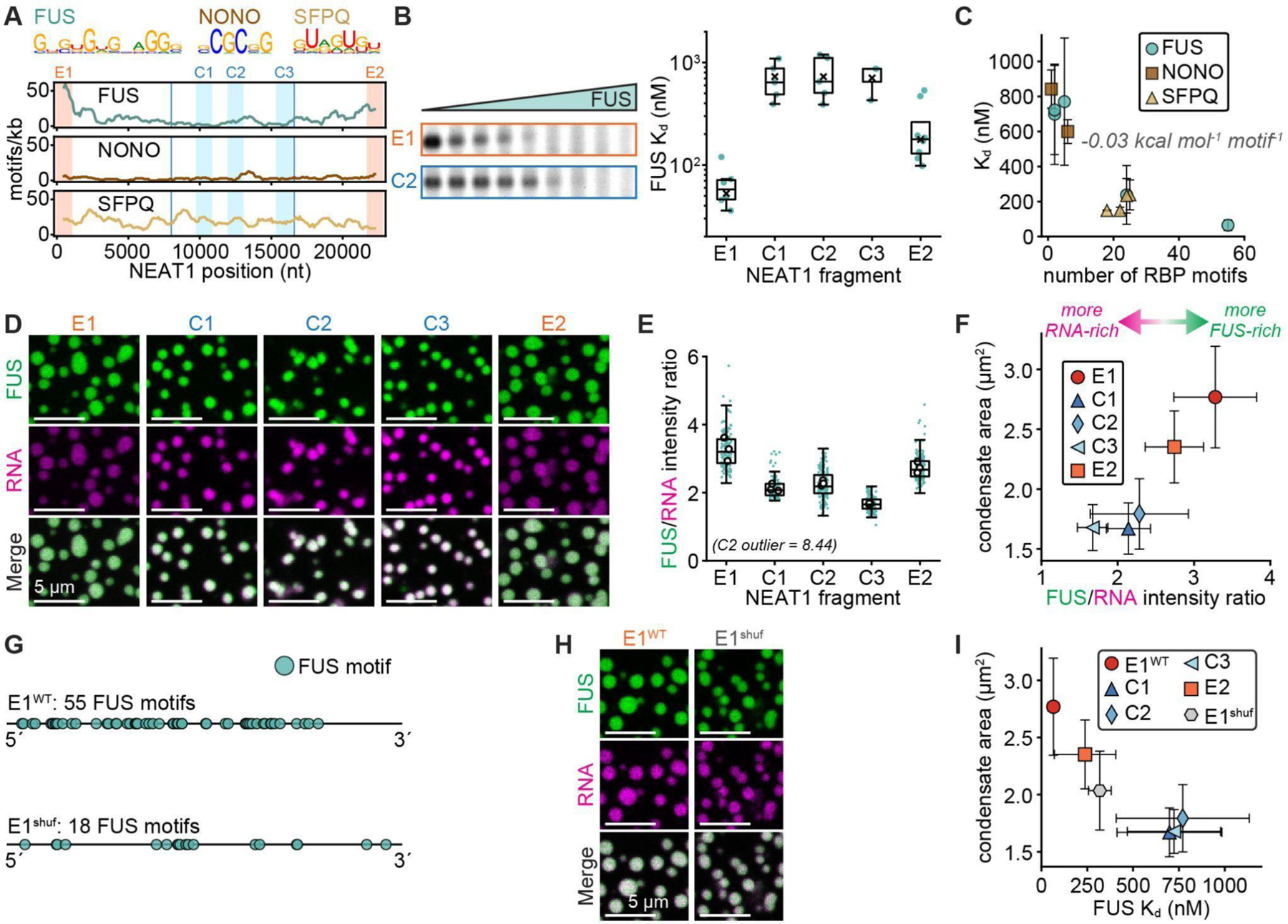
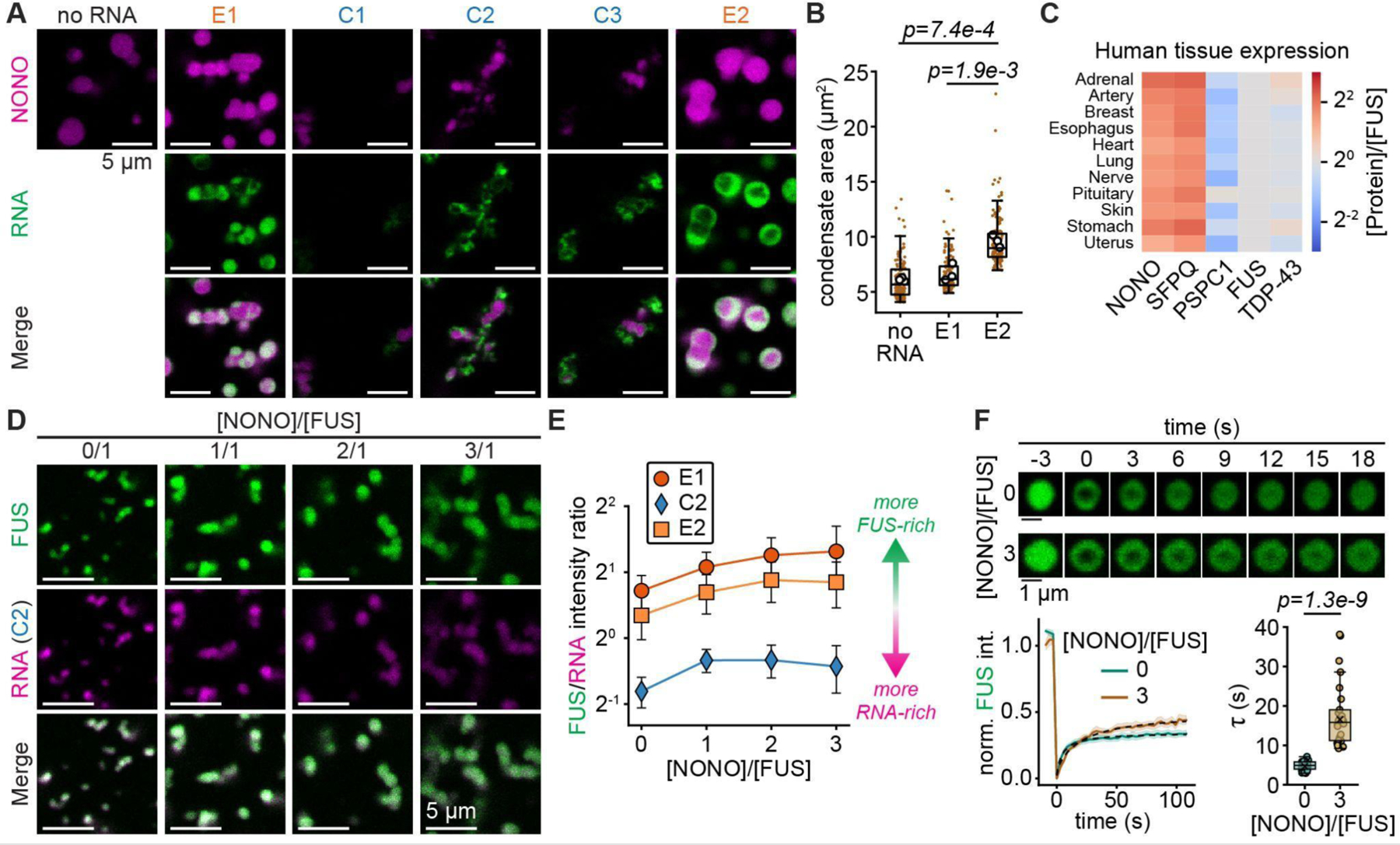
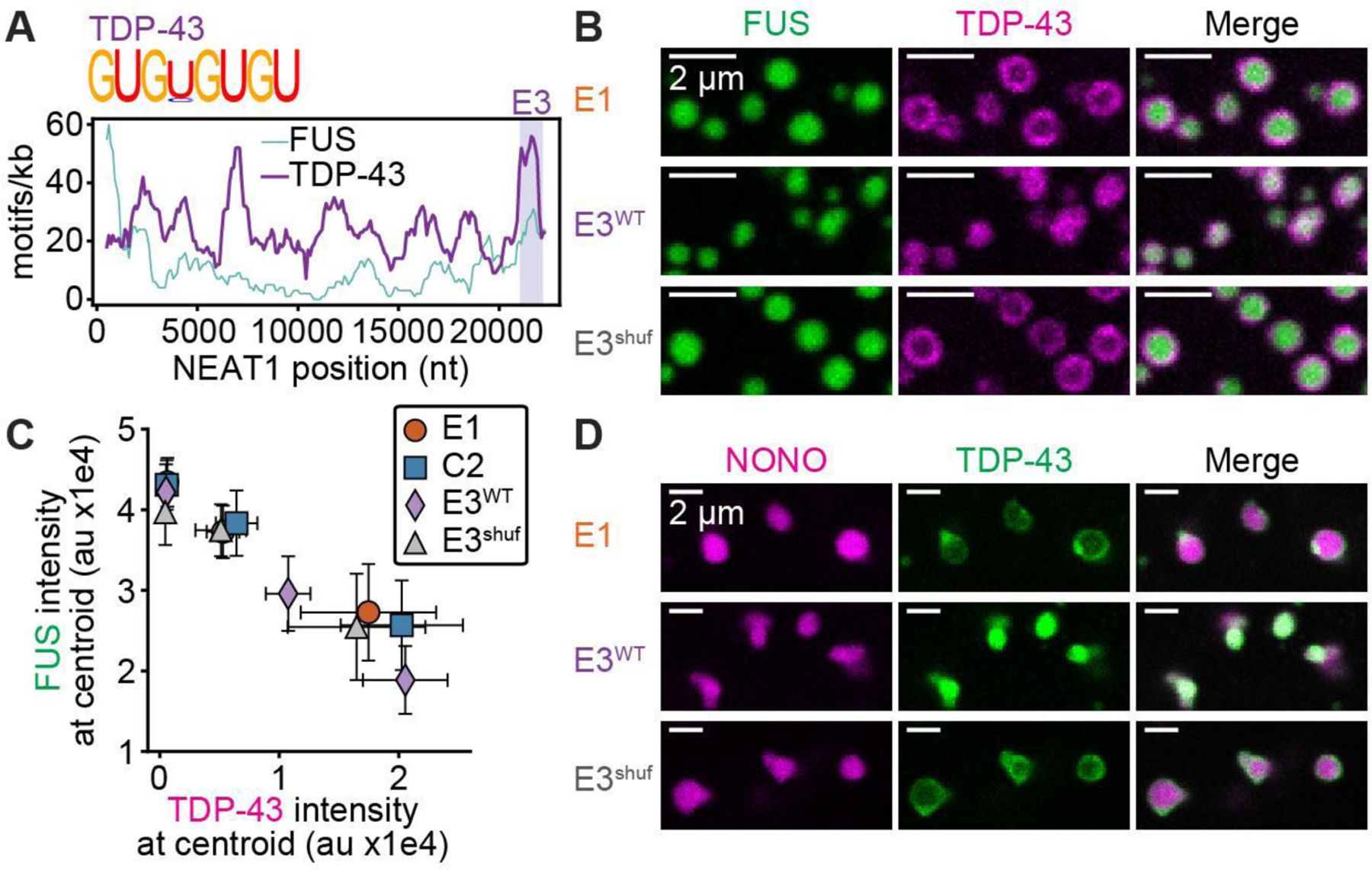
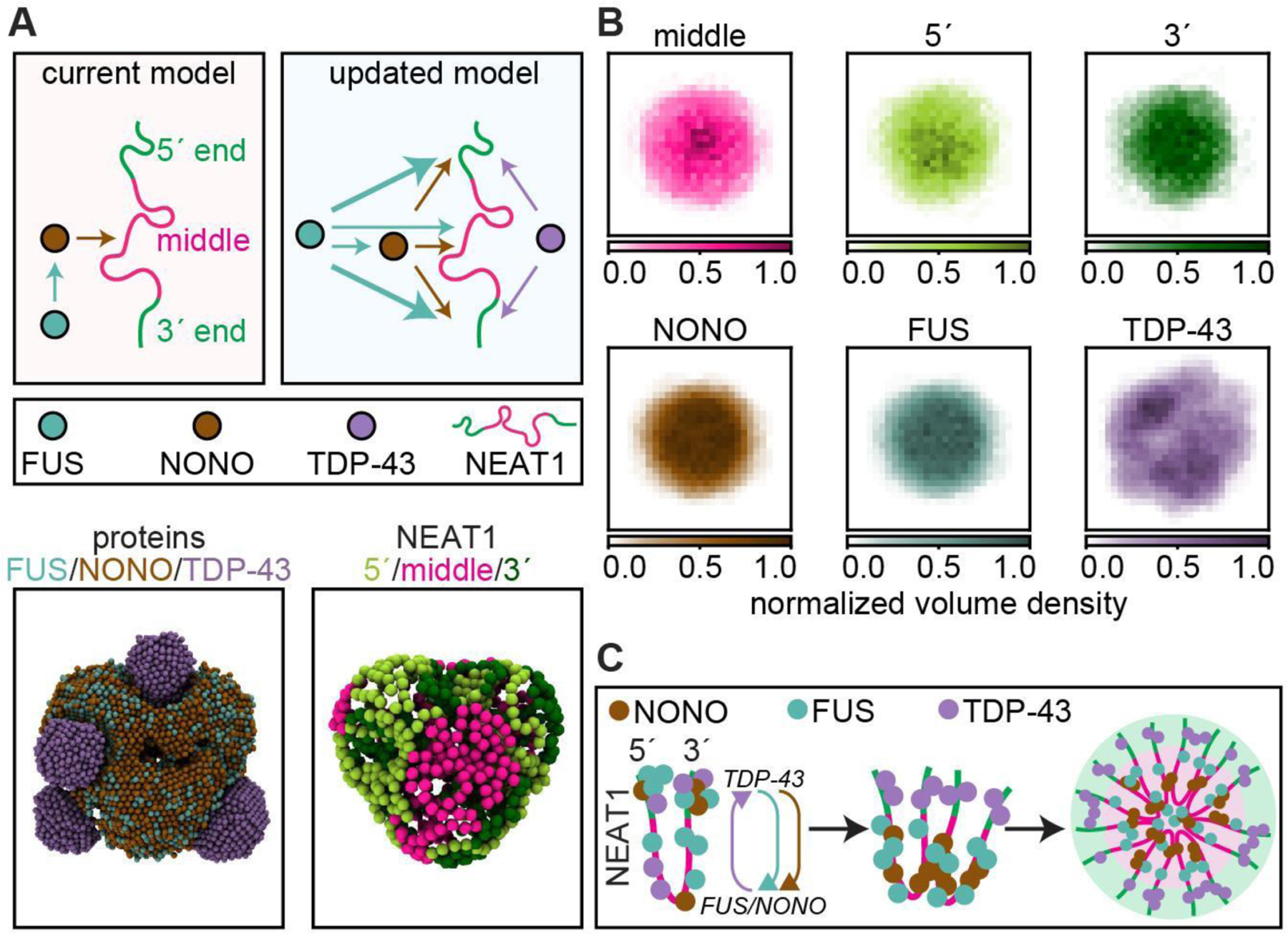
Biomolecular condensates mediate diverse and essential cellular functions by compartmentalizing biochemical pathways. Many condensates have internal subdomains with distinct compositional identities. A major challenge lies in dissecting the multicomponent logic that relates biomolecular features to emergent condensate organization. Nuclear paraspeckles are paradigmatic examples of multidomain condensates, comprising core and shell layers with distinct compositions that are scaffolded by the lncRNA NEAT1, which spans both layers. A prevailing model of paraspeckle assembly proposes that core proteins bind directly and specifically to core-associated NEAT1 domains. Combining informatics and biochemistry, we unexpectedly find that the essential core proteins FUS and NONO bind and condense preferentially with shell-associated NEAT1 domains. The shell protein TDP-43 exhibits similar NEAT1 domain preferences on its own but forms surfactant-like shell layers around core protein-driven condensates when both are present. Together, experiments and physics-based simulations suggest that competitive RNA binding and immiscibility between core and shell proteins order paraspeckle layers. More generally, we propose that subcondensate organization can spontaneously arise from a balance of collaborative and competitive protein binding to the same domains of a lncRNA.
Keywords: RNA; RNA-binding proteins; biomolecular condensates; nuclear paraspeckles.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
Update of
-
Immiscible proteins compete for RNA binding to order condensate layers.bioRxiv [Preprint]. 2025 Mar 19:2025.03.18.644007. doi: 10.1101/2025.03.18.644007. bioRxiv. 2025. Update in: Proc Natl Acad Sci U S A. 2025 Aug 12;122(32):e2504778122. doi: 10.1073/pnas.2504778122. PMID: 40166346 Free PMC article. Updated. Preprint.
Similar articles
-
Immiscible proteins compete for RNA binding to order condensate layers.bioRxiv [Preprint]. 2025 Mar 19:2025.03.18.644007. doi: 10.1101/2025.03.18.644007. bioRxiv. 2025. Update in: Proc Natl Acad Sci U S A. 2025 Aug 12;122(32):e2504778122. doi: 10.1073/pnas.2504778122. PMID: 40166346 Free PMC article. Updated. Preprint.
-
Regulation of physiological and pathological condensates by molecular chaperones.FEBS J. 2025 Jul;292(13):3271-3297. doi: 10.1111/febs.17390. Epub 2025 Jan 5. FEBS J. 2025. PMID: 39756021 Free PMC article. Review.
-
Client-scaffold interactions suppress aggregation of a client protein in model condensates.Proc Natl Acad Sci U S A. 2025 Aug 19;122(33):e2508403122. doi: 10.1073/pnas.2508403122. Epub 2025 Aug 12. Proc Natl Acad Sci U S A. 2025. PMID: 40794831
-
mTOR regulates aerobic glycolysis through NEAT1 and nuclear paraspeckle-mediated mechanism in hepatocellular carcinoma.Theranostics. 2022 Apr 24;12(7):3518-3533. doi: 10.7150/thno.72581. eCollection 2022. Theranostics. 2022. PMID: 35547764 Free PMC article.
-
RNA-associated nuclear condensates: Where the nucleus keeps its RNAs in check.Mol Cells. 2025 Aug;48(8):100240. doi: 10.1016/j.mocell.2025.100240. Epub 2025 Jun 15. Mol Cells. 2025. PMID: 40527409 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
- R35GM156800/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- T32GM153505/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- R01GM081506/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- R35 GM156800/GM/NIGMS NIH HHS/United States
- MCB 2044895/National Science Foundation (NSF)
LinkOut - more resources
Full Text Sources
