

Ex vivo- generated lymphoid progenitors encompass both T cell and innate lymphoid cell fates
- PMID: 40771806
- PMCID: PMC12325321
- DOI: 10.3389/fimmu.2025.1617707
Ex vivo- generated lymphoid progenitors encompass both T cell and innate lymphoid cell fates
Abstract
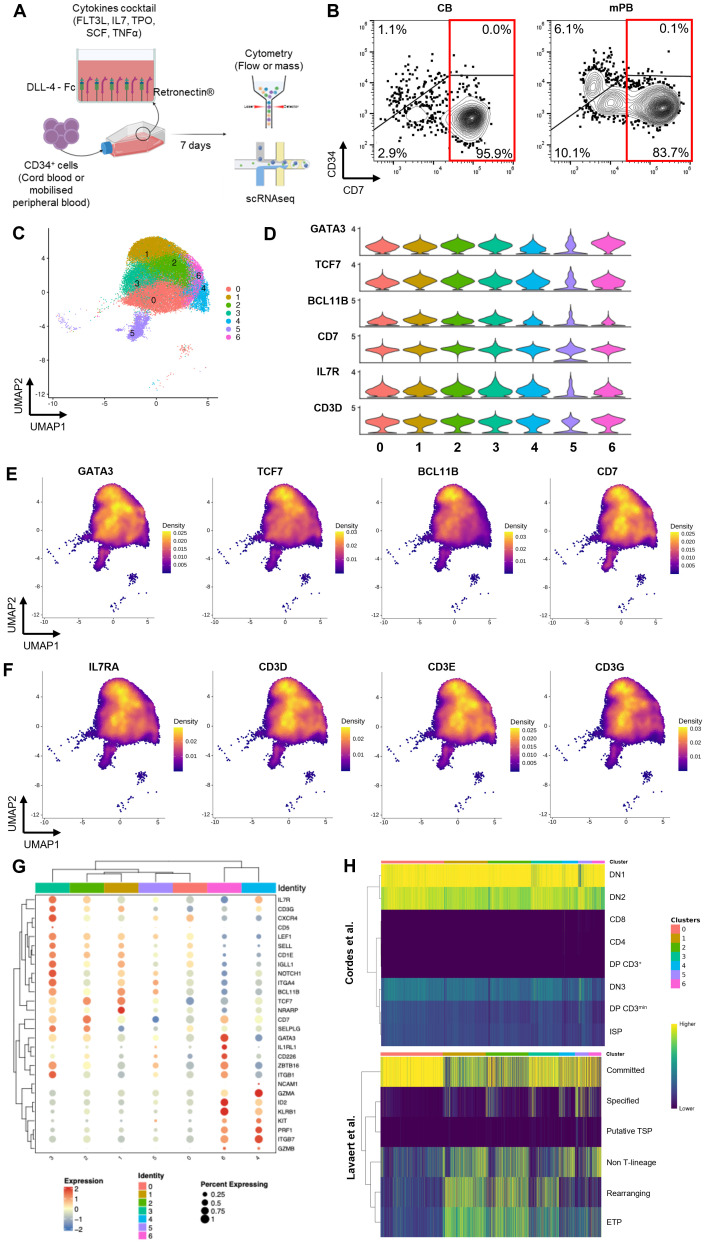
Introduction: We previously established a feeder-free cell therapy platform for the ex vivo generation of lymphoid-primed progenitors using immobilized Delta-like ligand 4 (DLL4). In vivo studies demonstrated that adoptive transfer of these progenitors accelerates T cell reconstitution following thymic engraftment.
Method: To further explore the full therapeutic potential of this cell product, we performed a comprehensive molecular and phenotypic characterization using single cell RNA sequencing and mass cytometry analysis.
Results: Our analysis revealed the presence of distinct cell subsets within the cellular product characterized mainly by commitment to lymphoid lineages. Using integrated transcriptomic analyses to compare these ex vivo-generated progenitors to in vivo human thymocytes, we revealed strong similarities with early stages of T cell development, underscoring the physiological relevance of our system. We also delineated two distinct developmental trajectories within the CD7+ progenitor population: a T cell-oriented path, marked by CD5 upregulation, and an innate lymphoid cell (ILC)-oriented branch, identified by CD161 expression and an ILC-like gene signature. Despite these lineage predispositions, both subsets demonstrated plasticity, retaining the ability to differentiate into both T cells and natural killer (NK) cells in vitro. Additionally, in our experimental setting, we observed that BCL11B, a transcription factor essential for T cell commitment, regulates negatively myeloid cell differentiation while preserving the potential for NK cell development.
Conclusion: These findings underscore the versatility of DLL4-based lymphoid progenitors in generating either T cells or ILCs in response to environmental cues. This research paves the way for innovative cell therapy approaches to treat immune deficiencies and cancer- and age-related immune dysfunctions.
Keywords: T cell progenitors; ex vivo differentiation; hematopoietic stem cell; lymphoid cell development; scRNAseq.
Copyright © 2025 Gaudeaux, Paillet, Abou Alezz, Moirangthem, Cascione, Martin Corredera, Dolens, De Mulder, Velghe, Vandekerckhove, Lavaert, Robil, Corneau, Sadek, Rault, Joshi, de la Grange, Staal, Taghon, Negre, Ditadi, André and Soheili.
Conflict of interest statement
The Human Lymphohematopoiesis Laboratory and Smart Immune have contracted an industrial collaboration agreement to conduct this project. PG, is a former employee of Smart Immune. JP, RDM, MMC, HS, PR, ON, T-SS are employees of Smart Immune. IA is a co-founder and owns shares of Smart Immune. IA, T-SS, RDM are inventors of patents related to ProTcell production WO2016/055396 and WO2018/146297 which are licensed to Smart Immune. PdlG is an employee and co-founder of GenoSplice. NR is an employee of GenoSplice. FJTS is a scientific advisor for Genewity. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Combined transient ablation and single-cell RNA-sequencing reveals the development of medullary thymic epithelial cells.Elife. 2020 Nov 23;9:e60188. doi: 10.7554/eLife.60188. Elife. 2020. PMID: 33226342 Free PMC article.
-
A feedback amplifier circuit with Notch and E2A orchestrates T-cell fate and suppresses the innate lymphoid cell lineages during thymic ontogeny.Genes Dev. 2025 Mar 3;39(5-6):384-400. doi: 10.1101/gad.352111.124. Genes Dev. 2025. PMID: 39904558 Free PMC article.
-
Targeting BCL11B in CAR-engineered lymphoid progenitors drives NK-like cell development with prolonged anti-leukemic activity.Mol Ther. 2025 Apr 2;33(4):1584-1607. doi: 10.1016/j.ymthe.2025.02.024. Epub 2025 Feb 15. Mol Ther. 2025. PMID: 39955618 Free PMC article.
-
A new model of human lymphopoiesis across development and aging.Trends Immunol. 2024 Jul;45(7):495-510. doi: 10.1016/j.it.2024.05.007. Epub 2024 Jun 21. Trends Immunol. 2024. PMID: 38908962 Review.
-
Impact of residual disease as a prognostic factor for survival in women with advanced epithelial ovarian cancer after primary surgery.Cochrane Database Syst Rev. 2022 Sep 26;9(9):CD015048. doi: 10.1002/14651858.CD015048.pub2. Cochrane Database Syst Rev. 2022. PMID: 36161421 Free PMC article.
References
-
- Bartelink IH, Belitser SV, Knibbe CA, Danhof M, de Pagter AJ, Egberts TC, et al. Immune reconstitution kinetics as an early predictor for mortality using various hematopoietic stem cell sources in children. Biol Blood Marrow Transplant. (2013) 19:305–13. doi: 10.1016/j.bbmt.2012.10.010, PMID: - DOI - PubMed
-
- Clave E, Lisini D, Douay C, Giorgiani G, Busson M, Zecca M, et al. Thymic function recovery after unrelated donor cord blood or T-cell depleted HLA-haploidentical stem cell transplantation correlates with leukemia relapse. Front Immunol. (2013) 4:54. doi: 10.3389/fimmu.2013.00054, PMID: - DOI - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources