

Structure of the T9SS PorKN ring complex reveals conformational plasticity based on the repurposed FGE fold
- PMID: 40772760
- PMCID: PMC12421860
- DOI: 10.1128/mbio.01799-25
Structure of the T9SS PorKN ring complex reveals conformational plasticity based on the repurposed FGE fold
Abstract
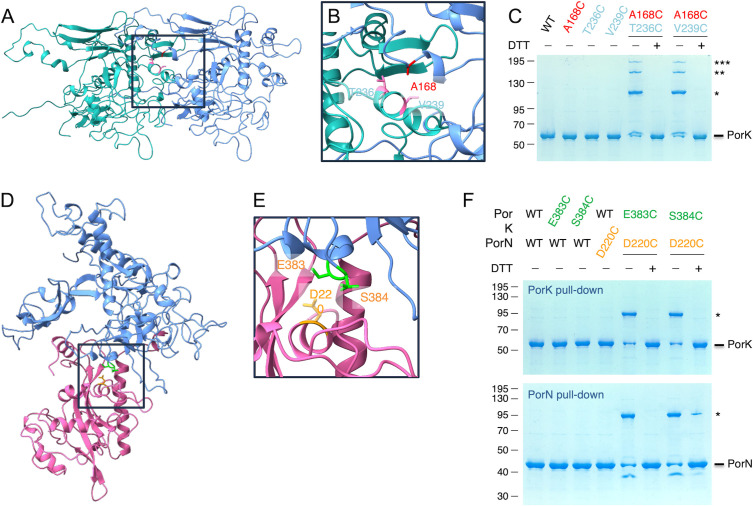
The type IX secretion system (T9SS) is a protein secretion machinery unique to the Bacteroidetes-Chlorobi-Fibrobacteres superphylum, which plays crucial roles in bacterial pathogenesis and gliding motility. It is composed of >15 proteins, including the proton-motive force-dependent PorLM motor, the PorKN ring anchored to the outer membrane, and the Sov translocon. Here, we present the cryo-electron microscopy (EM) structure of the PorKN ring complex from Porphyromonas gingivalis at 3.2 Å resolution. Our structural analysis reveals that PorK contains a repurposed formylglycine-generating enzyme-like fold, which serves as a structural hub for complex assembly rather than enzymatic activity. The complex exhibits a 33-fold symmetry with PorK and PorN assembling two tightly packed and wedged subrings. The structure reveals previously uncharacterized N- and C-terminal helices in PorN that are crucial for PorK binding and complex stability. By combining our high-resolution structure with in situ cryo-electron tomography data, we propose a mechanism whereby PorKN undergoes conformational changes during substrate transport, transitioning between 50° and 90° states relative to the membrane plane. Finally, structural predictions coupled to site-directed disulfide cross-linking identified contacts between PorM and the PorKN ring. Collectively, these findings provide crucial insights into the molecular architecture and dynamic behavior of the T9SS machinery, advancing our understanding of bacterial protein secretion mechanisms.IMPORTANCEThe bacterial type IX secretion system (T9SS) is essential for processes such as gliding motility and secretion of virulence factors. In Porphyromonas gingivalis, a major periodontal pathogen, the T9SS transports over 30 virulence-associated proteins, making it central to disease development. The T9SS core is composed of PorLM motors that are thought to energize the PorKN outer membrane-associated ring. However, the molecular architecture of the PorKN ring has remained unresolved. Here, we present its atomic-resolution cryo-EM structure, revealing a formylglycine-generating enzyme-like fold in PorK that mediates PorK-PorN interactions through specific insertion motifs. Our results show that the ring exhibits intrinsic structural plasticity, including dynamic flexibility and variable stoichiometry. AlphaFold models and disulfide cross-linking experiments further provide information on how PorLM motors are connected to the PorKN ring. These insights redefine our understanding of the T9SS mechanism of action and offer a structural framework for the development of targeted antimicrobial strategies.
Keywords: cryo-EM; surface structures; type 9 protein secretion.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
Insights into type IX secretion from PorKN cogwheel structure bound to PorG and attachment complexes.Nat Commun. 2025 Aug 19;16(1):7735. doi: 10.1038/s41467-025-63163-1. Nat Commun. 2025. PMID: 40830366 Free PMC article.
-
A molecular conveyor belt-associated protein controls the rotational direction of the bacterial type 9 secretion system.mBio. 2025 Jul 9;16(7):e0112525. doi: 10.1128/mbio.01125-25. Epub 2025 Jun 13. mBio. 2025. PMID: 40511966 Free PMC article.
-
Protein Interactome Analysis of the Type IX Secretion System Identifies PorW as the Missing Link between the PorK/N Ring Complex and the Sov Translocon.Microbiol Spectr. 2022 Feb 23;10(1):e0160221. doi: 10.1128/spectrum.01602-21. Epub 2022 Jan 12. Microbiol Spectr. 2022. PMID: 35019767 Free PMC article.
-
Type IX secretion: the generation of bacterial cell surface coatings involved in virulence, gliding motility and the degradation of complex biopolymers.Mol Microbiol. 2017 Oct;106(1):35-53. doi: 10.1111/mmi.13752. Epub 2017 Aug 9. Mol Microbiol. 2017. PMID: 28714554 Review.
-
Aspects of Genetic Diversity, Host Specificity and Public Health Significance of Single-Celled Intestinal Parasites Commonly Observed in Humans and Mostly Referred to as 'Non-Pathogenic'.APMIS. 2025 Sep;133(9):e70036. doi: 10.1111/apm.70036. APMIS. 2025. PMID: 40923351 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
